
形象锦言:美丽国土 古茶原乡
推荐阅读:中国国土经济学会研究室“深呼吸古茶原乡”一、二号联合观察坐标点(三都都江镇-水龙乡)
栏目编研:芬儿、后卿、新宇
贵州茶树种质资源遗传多样性、群体结构和遗传分化研究
安红卫, 宋勤飞, 牛素贞
(贵州大学 茶学院, 贵州 贵阳 550025)
茶是世界上三大饮料中有益人体健康的饮品[1-2],具有很高的营养和药用价值。茶的丰富风味由近700种生物活性化合物组成,如儿茶素、茶氨酸、咖啡因和挥发物[3-4]。茶树[Camelliasinensis(L.)O. Kuntze],原产中国西南地区,而贵州位于茶树原产地的中心地带[5-6],该地区独特的地貌特征、多样的气候、丰富的降水及茶树的异花授粉特性,保存了丰富的野生茶树、古老地方品种和具有不同形态特征的现代地方品种[7],因此对贵州境内茶树种质资源的分布状况及遗传多样性的研究具有重要意义。本研究采用EST-SSR分子标记,对贵州省境内的144份茶树种质资源进行遗传多样性、群体结构及遗传分化分析,探索贵州茶树种质资源的分布规律及保护与利用方向。
1 材料与方法
1.1 材料
以144份贵州茶树种质资源为材料(表1)。参考陈亮[8]提出的茶树分类系统,144份茶树种质资源包含有14个秃房茶(C.gymnogyna),80个茶[C.sinensis(L.) O. Kuntze],39个大厂茶[C.tachangensis(F.C.Zhang)],11个疑似大理茶[C.taliensis(W.W.Smith)];海拔500~1 000 m有23个,海拔1 000~1 300 m有59个,海拔1 300~1 500 m有34个,海拔1 500 m以上有28个;根据地区分类则黔中地区11个,黔北地区36个,黔南地区27个,黔东南地区18个,黔西南地区52个。
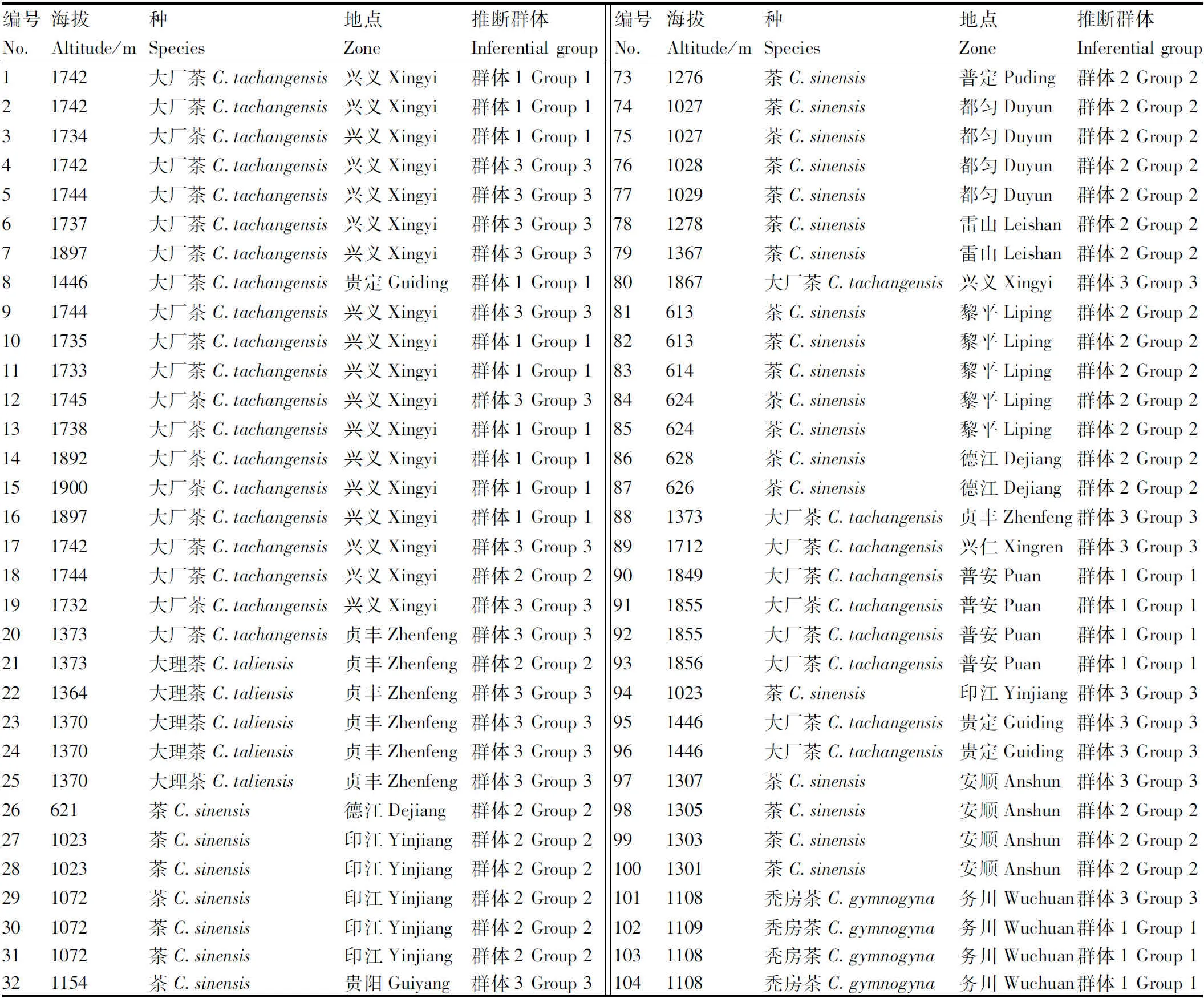
表1 一百四十四份贵州茶树种质资源信息
1.2 试验方法
1.2.1茶树基因组DNA提取及EST-SSR引物的选择
采用改进的CTAB法[9]提取144个茶树种质资源基因组DNA。所用引物(表2)参考乔婷婷等[10-11]研究中的引物,从中共筛选出68条多态性好、条带清晰的引物开展研究。
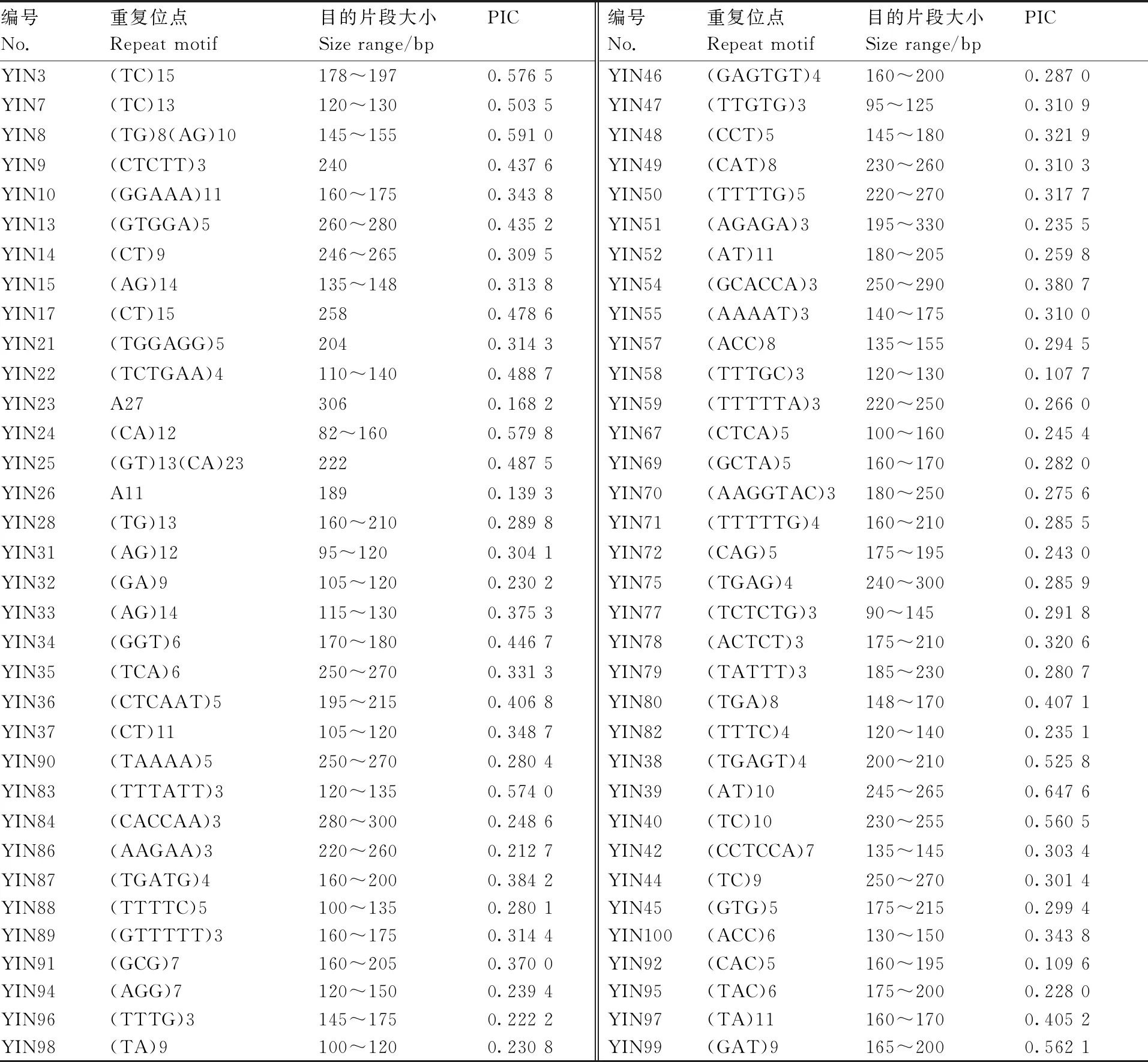
表2 SSR引物信息
1.2.2 EST-SSR扩增技术体系及产物检测
参考刘本英[12]的方法进行PCR扩增,在DYY-6C电泳仪和垂直电泳槽DYCZ-30上进行凝胶电泳,120 V电压下电泳120 min,电泳后银染显色。
1.2.3 数据处理及分析
将清晰条带记为“1”,无带或不易分辨的弱带记为“0”,建立0、1数据库。使用Popgene(version 1.31)计算有效等位基因数(Ne)、Nei’s遗传多样性(H)、Shannon信息指数(I)、遗传分化程度(Ht)、群体内基因多样性(Hs)、群体间遗传分化系数(Gst)、基因流(Nm)等信息;使用Structure分析种群结构,根据daltaK和LNP(K)确定群体K的数量,连续运行K=1~9估计重复之间的方差,确定最优种群数量。每次运行的Burn-in period设置为30 000,重复100 000次,每个K值重复计算3次;使用DARWIN制作主成分分析图和聚类分析图。
2 结果与分析
2.1 遗传多样性分析
通过对144份茶树种质资源的遗传多样性分析,结果显示(表3),4个种中茶的多态性位点百分率和Shannon’s 指数最高,秃房茶的有效等位基因数和Nei’s遗传多样性最高,疑似大理茶多态性位点百分率、有效等位基因数、Shannon’s 指数以及Nei’s遗传多样性最低;5个地区中黔北的有效等位基因数、Nei’s遗传多样性、Shannon’s 指数最高,其次是黔西南的Nei’s遗传多样性和Shannon’s较高,黔中的有效等位基因数、Nei’s遗传多样性、Shannon’s 指数最低;4个海拔范围中1 000~1 300 m的有效等位基因数、Shannon’s 指数最高,海拔范围1 300~1 500 m的Nei’s遗传多样性最高,海拔1 500 m以上的有效等位基因数、Nei’s遗传多样性、Shannon’s 指数最低。
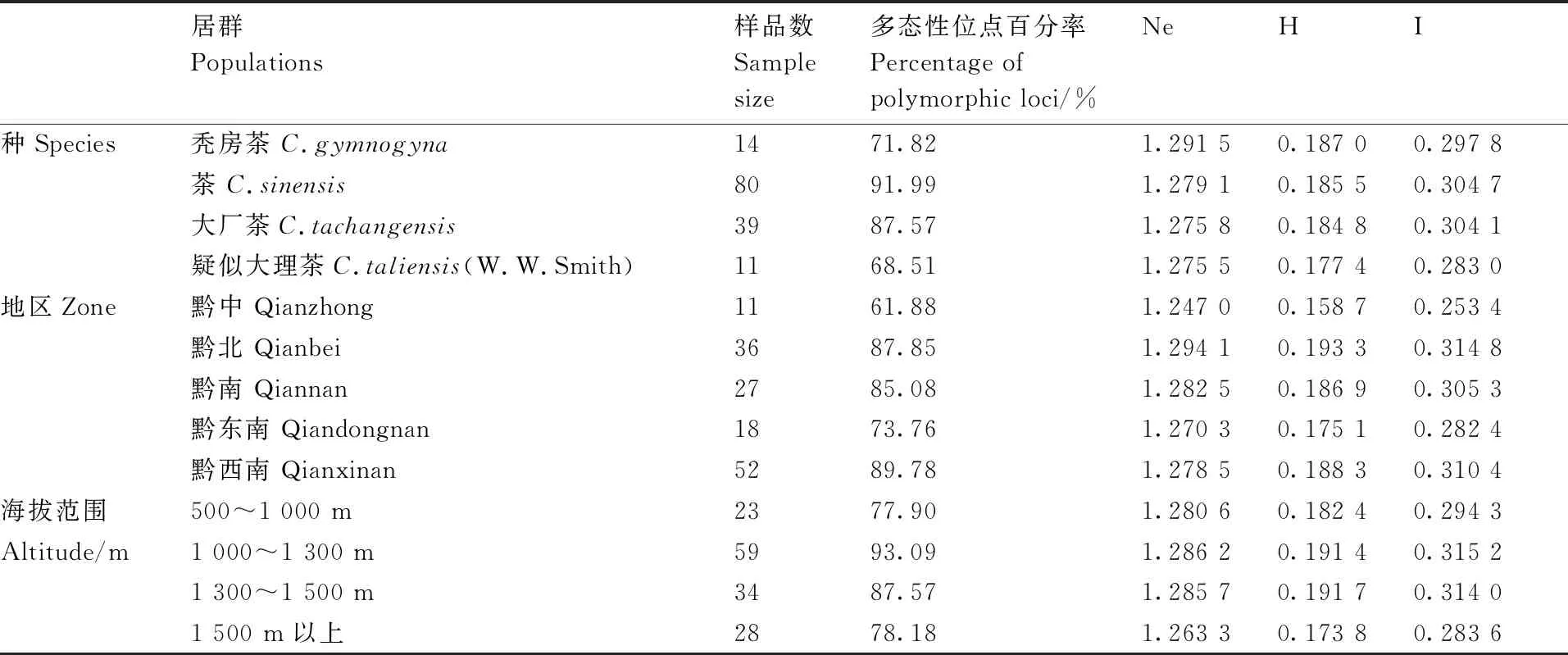
表3 茶树种质资源的遗传多样性分析
2.2 群体结构分析
通过对144份贵州茶树种质资源的群体结构分析,结果显示(图1-A),随着K值的增加,MeanlnP(K)值也持续增加,且当K=2时,ΔK值最大,出现明显的峰值,表明144份茶树种质资源有两个祖先(图1-B)。遗传组分(Q值)≥0.8[13]表示该材料的遗传背景相对比较单一,遗传组分(Q值)<0.8属于混合祖先,因此,本研究的群体被分为2个祖先组和1个混合祖先组,分别被命名为Gro1、Gro2、Gro3。
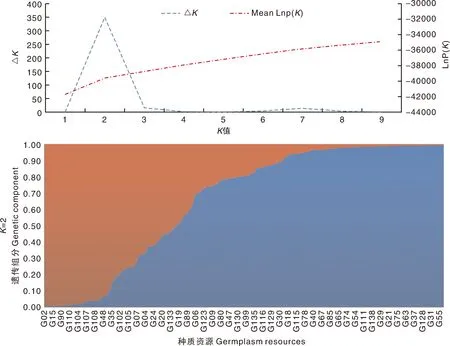
图1 一百四十四份茶树种质资源遗传结构分析
Gro1共28份材料(图2-A),根据种包含19份大厂茶(67.86%)、5份秃房茶(17.86%)、4份疑似大理茶(14.29%);根据海拔范围包括海拔1 500 m以上有17份(60.71%)、海拔范围500~1 000 m有2份(7.14%)、海拔范围1 000~1 300 m有5份(17.86%)、海拔范围1 300~1 500 m有4份(14.29%);根据地区包含黔西南材料17份(60.71%)、黔北材料7份(25.00%)、黔南材料4份(14.29%)。Gro2共76份材料(图2-B),根据种包含70份茶(92.11%)、3份秃房茶(3.95%),2份疑似大理茶(2.63%);根据海拔范围包括海拔1 000~1 300 m有41份(53.95%)、海拔范围500~1 000 m有18份(23.68%)、海拔范围1 300~1 500 m有16份(21.05%)、海拔范围1 500 m以上有1份(1.32%);根据地区包含黔北材料19份(25.00%)、黔南材料16份(21.05%)、黔东南材料16份(21.05%)、黔西南材料16份(25.00%)、黔中材料9份(11.84%)。Gro3共40份材料(图2-C),根据种包含19份大厂茶(47.50%)、10份茶(25.00%)、6份秃房茶(15.00%)、5份疑似大理茶(12.50%);根据海拔范围包括海拔范围500~1 000 m有3份(7.50%)、海拔范围1 000~1 300 m有13份(32.50%)、海拔范围1 300~1 500 m有14份(35.00%)、海拔范围1 500 m以上有10份(25.00%);根据地区包含黔西南材料19份(47.50%)、黔北材料10份(25.00%)、黔南材料7份(17.50%)、黔东南材料2份(5.00%)、黔中材料2份(5.00%)。主成分分析结果显示(图3),Gro1分布在纵坐标的右边,Gro2分布在纵坐标的左边,Gro3分散在纵坐标中间,进一步验证了Structure群体结构分析的准确性。NJ聚类分析结果显示,尽管部分种质资源被分到其他组,大部分的材料聚类结果(图4)与Structure群体结构的分析结果基本一致。
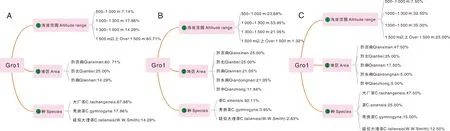
A、B、C分别为Gro1、Gro2、Gro3。
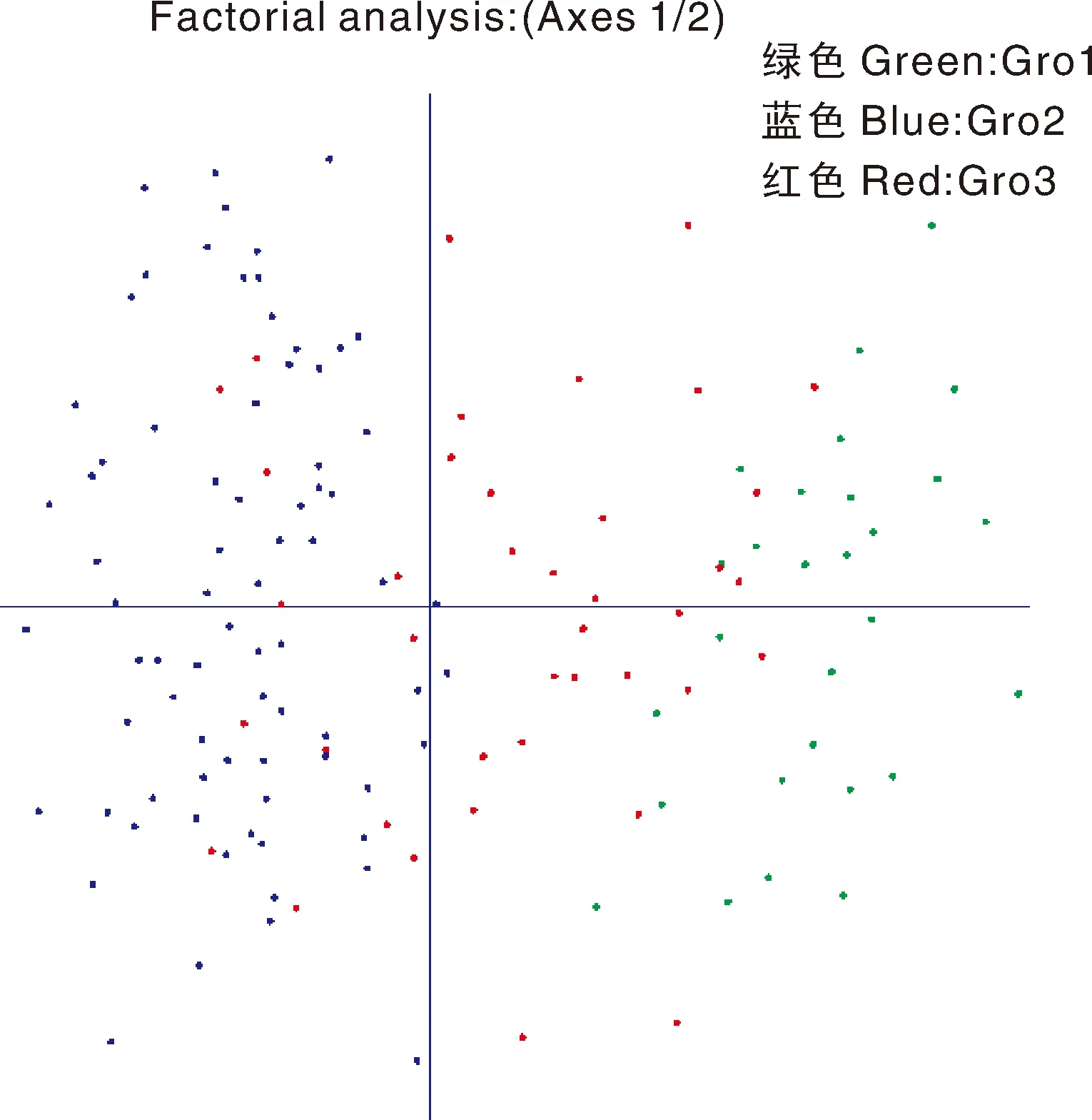
图3 一百四十四份茶树种质资源主成分分析图

A. 绿色,Gro1;蓝色,Gro2;红色,Gro3。B. 红色,秃房茶;紫色,茶;绿色,大厂茶;蓝色,疑似大理茶。C. 蓝色,500~1 000 m;红色,1 000~1 300 m;紫色,1 300~1 500 m;绿色,1 500 m以上。
2.3 遗传距离分析
通过对144份贵州茶树种质资源的遗传距离分析,结果显示(表4),4个种间的遗传距离变化范围为0.017 8~0.038 9,其中秃房茶群体与疑似大理茶群体的遗传距离最大,其次秃房茶群体与茶群体间的遗传距离较大,茶群体与大厂茶群体的遗传距离最小;地区间的遗传距离分析结果显示(表5),5个分布地区的遗传距离范围为0.007 8~0.032 4,其中黔西南与黔中的遗传距离最大,其次为黔南和黔中,黔南和黔北的遗传距离最小;不同海拔间遗传距离分析结果显示(表6),4个海拔范围的遗传距离范围为0.006 6~0.025 6,海拔范围在1 500 m以上和500~1 000 m的遗传距离最大,其次为1 500 m和1 000~1 300 m,海拔范围1 000~1 300 m和1 300~1 500 m的遗传距离最小。

表4 不同茶树种质资源的遗传距离

表5 不同地区茶树种质资源的遗传距离
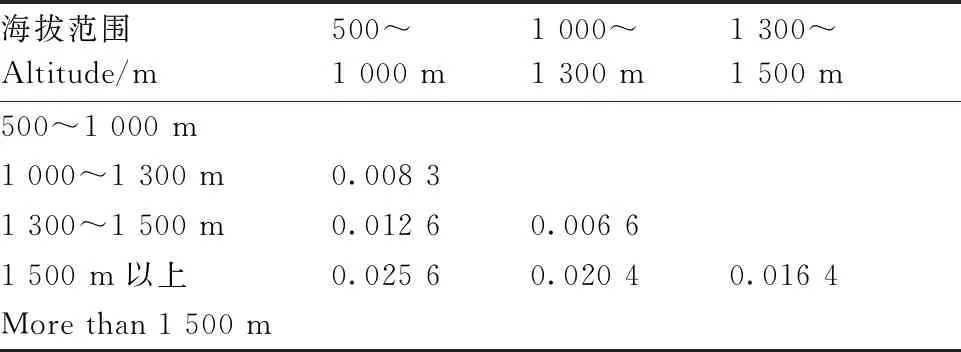
表6 不同海拔范围茶树种质资源的遗传距离
2.4 遗传分化分析
通过对144份贵州茶树种质资源进行遗传分化分析,结果表明(表7),遗传分化程度Ht值在种间最高为0.199 6,在地区间最低为0.193 6;群体内基因多样性Hs值在海拔间最高为0.184 8,在地区间最低为0.180 5;遗传分化系数Gst值在种间最高为0.079 7,在海拔间最低为0.047 0,说明遗传变异主要来自于群体内的个体间;基因流Nm值在海拔间最高为10.134 8,其次是地区间为6.885 2,种间基因流Nm值最低为5.774 1。说明贵州不同海拔的茶树种质资源间基因交流最为频繁,其次是不同地区间,种间相对较小。
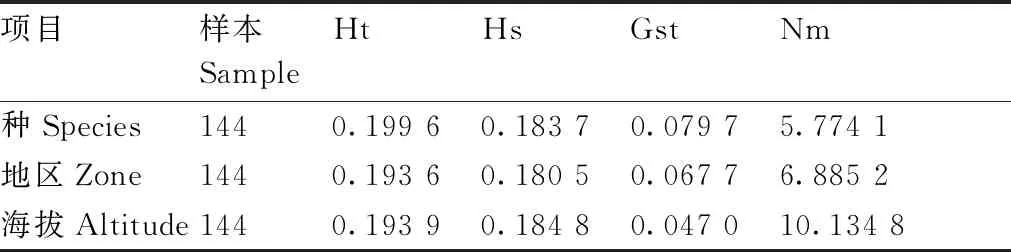
表7 茶树种质资源的遗传分化分析
3 讨论
3.1 遗传多样性分析
本研究用EST-SSR引物研究了贵州境内的茶树种质资源,引物的平均多态信息为0.338 4,属于中等水平[14],10对引物的多态性较高(PIC>0.5),55对引物的多态性适中(0.2
3.2 群体结构分析
张明泽等[18-20]的研究结果显示,茶树不是按地理来源进行聚类。本研究将144份茶树种质资源按海拔范围及种都可以分为3类,Gro1主要是分布海拔1 500 m以上的大厂茶群体,Gro2主要是分布在海拔范围1 000~1 300 m的茶群体。对材料的来源地分析结果显示,与材料来源地没有明显的相关性,这与前人的研究结果一致。大厂茶和秃房茶被聚为一类(图3-B),可能大厂茶和秃房茶都是属于野生型茶树有关系,而栽培型茶被分为另外一个群体。本研究中对4个种的海拔分布分析结果显示,大厂茶主要分布在海拔1 300 m以上,茶分布在海拔1 500 m以下,秃房茶主要分布在海拔1 300 m以下。本研究的群体结构分析结果对贵州茶树品种的选育及栽培具有一定的指导意义。
3.3 遗传距离分析
陈亮[21]研究认为,原始茶组植物演化线路分两条线路,一条是大厂茶向秃房茶方向演化,另一条为疑似大理茶向茶方向演化。本研究对144份贵州茶树种质资源按种进行遗传距离分析结果显示,大厂茶与秃房茶遗传距离最小,亲缘关系最近,秃房茶与疑似大理茶及茶的遗传距离较大,亲缘关系较远,印证了第一条演化路线。但本研究发现茶和大厂茶的遗传距离最小,亲缘关系最近,茶和疑似大理茶的遗传距离较小,亲缘关系较近,与该研究的第二条演化路线结果不符,这可能与本研究中疑似大理茶的数量相对较少有关系。
贵州姑菁野生茶树形态多样性及相关性研究显示[22],茶树叶、果器官与海拔范围呈极显著相关并表现出差异性,杨亚军等[23-24]认为,云贵高原可能为茶树的生态演化区, 其分布、演化受气候条件影响,贵州野生茶树资源地理分布和形态特征与气候要素的关系[25]中认为气候影响了贵州茶树资源的分布、演化及部分形态特征。本研究发现,茶树种质资源的海拔范围差距越大,遗传距离越大,亲缘关系越远,即遗传距离与海拔范围呈正相关关系,本研究进一步说明贵州野生茶树种质资源的分布可能与海拔有关,为更进一步发现贵州野生茶树种质资源的演化和分布规律奠定基础。
3.4 遗传分化分析
估算群体间的遗传分化系数可区分群体间和群体内相对遗传变异大小,是解释群体遗传变异程度的主要依据[26]。群体多样性水平或分化程度差异的可能原因在于选择、基因流、隔离或随机漂变等。Curnow等[27]认为Nm大于1,则表明基因流是影响群体遗传分化的主要原因,基因流动可以消除因地理或生殖隔离导致的不同地方群体甚至是物种间的遗传差异,减小种间或种内群体间的分化程度,进一步提高群体或物种的多样性和适应力。有研究结果表明,西双版纳[28]不同地理区域的古茶树基因流Nm为1.624,城步峒茶[29]不同地理群体间的基因流Nm为7.01,南江大叶茶树[30]不同分布区域群体间基因流Nm为3.554 9,前人的研究表明,茶树群体之间存在较强的基因流。本试验研究结果显示,贵州野生茶树遗传变异主要来自于群体内的个体间,群体间遗传分化系数与基因流之间呈现负相关,在不同海拔群体间的基因流值Nm为10.134 8、不同地区群体间的基因流Nm为6.885 2、种间的基因流Nm为5.774 1,说明贵州茶树群体之间存在较强的基因流,基因流影响了茶树群体的遗传结构,即较高的遗传多样性主要存在于茶树群体内,茶树群体间的遗传分化较低。这种遗传变异可能与茶的传播方式有关。茶树异花授粉,花期从6月初持续到12月末,花粉小,以虫媒为主,可以进行长距离传播。贵州地势由西向东倾斜,夏秋季节盛行西南季风及西风,也有利于茶树花粉的长距离传播,这是茶树长期进化适应生态环境的结果。同时贵州山多地少,这里的人民顺应山势地形建造房屋,开展日常生活以及进行茶的贸易活动,都有助于茶树种子的长距离传播。茶长期适应生态环境及人类活动的共同结果导致茶树群体间存在较大基因流,减弱了群体间的分化。