
��ˮ����Ұӿ����������֡�������ܶ����Ա仯*
��������ѧ����־ 2024��1��
���ߣ��� ��,�� ΰ,�Զ���,������,��ԪԪ,������,�����,�´���
(1:������ѧ������ѧѧԺ,����ʡ����������ɫ������Դ�������������ù��̼����о�����,�人 430056) (2:������ѧ,�־����ж���Ⱦ�ﻷ���뽡��Σ������ʡ�ص�ʵ����,�人 430056) (3:�й�ˮ����ѧ�о�Ժ����ˮ���о���,�人 430223)
��ͳ����������Բ����Ҫ����Margalef�ḻ��ָ����Shannon-Wienerָ����Pielous���ȶ�ָ����,������Щ���ֶ�����ָ��Ӧ�ù㷺,����ʵ��Ӧ���д���һЩ����:���Ƕ��������ֵ�ͬ�Դ�,δ�������ּ�Ĺ��ܲ����Լ����ǵķ����ϵ[1]��Խ��Խ����о��߿�ʼ��ע������ܶ�����[2-3]����������Լ��ڶ����Բ���п����ּ���Ե��ϵ,�������������ָ�����������ָ����,Ⱥ������ַḻ�Ⱥ;��ȶ���ͬ,�����������Կ��ܲ�ͬ[4]�����ܶ����Լ�Ⱥ�����ּ书�������Ķ�����,�������ܷḻ�ȡ����ܾ��ȶȺ�����ɢ�ȵ�[5]��������ͬ��������Ⱥ��,�����ּ���̬����������̬����״�IJ���,�����ڹ��ܶ����Է�����ڽϴ���졣�չ�����[6]�����˽�ɳ��������ˮ��վ��ӽ���ǰ������Ⱥ��������,��ϲ�ͬ�������״�IJ���,�Աȷ����˽���ǰ������������Ⱥ��ṹ���ܶ����Եı仯,���ִ�ӽ���ǰ������Ⱥ�����ϴ�,���̽�������ͽϴ���λ��ʳ��խ������Ӱ��ϴ��봫ͳ�����ֶ��������,��������Ժ��ܶ����Ը��ܷ�ӳ�����ֵĹ��ܸ�����,�Ի���в�Ȼ��Ŷ�������,���ľ�Ӧ��ǰ���Ķ����Բ�ȷ���[7-8]��Ŀǰ���ڶ������������Ե��о���Ҫ�����ں�������[9-11],�Ե�ˮ������о���Ȼ����[12-14];������ܶ����Ե��о�Ҳ�����ܵ�����[15-17]��
��ɳ����������������Դ�ḻ,�dz�����������Ա������ص�����[18-19]����Ұ�ˮ��վ��Ϫ���ˮ��վ�ǽ�ɳ�������������ܹ�ע��ˮ�繤��,�ֱ���2012��10�º�2013��5�¿�ʼ��ˮ���С�ˮ�繤�̽���������������,�ı���������Ϣ����,�Ը�������������Բ�����������в[20-21]��Ŀǰ����Ұӿ���������о�����,Gao��[22]�о�����Ұ�ˮ��վ����������������Ⱥ����������Ӱ��,������[23]�����õ�[24]�ֱ𱨵�����ˮ����Ұӿ����罭����Ϫ��������Ⱥ��ṹ�ı仯,��Щ�о���Ҫ��ע�ֲ����ε����,����������̬��Ⱥ�����ֶ����Է��档�����о��߶�,��ϲ�ͬ��������Բ�ȷ����ӿռ��ʱ���Ͽ�չ�Ա��о�,������������̽���ݼ������Խ�ɳ����������Ⱥ���Ӱ��[18]��
���о��������֡�������ܶ����Եȶ�����������Բ�ȷ���,̽�ֽ�ɳ��������Ұӿ�����������Ե�ʱ�ո��,��Ҫ�о�Ŀ�����:(1)��ˮ����Ұӿ�����ͬ����(���ס����С���β)�������ֶ����ԡ���������Ժ��ܶ����ԵĿռ�ֲ�;(2)��ˮǰ��ͬʱ����Ұӿ����������ֶ����ԡ���������Ժ��ܶ����Եı仯���ơ��о���������������˽�ˮ�繤�̽���Խ�ɳ��������������Ե�Ӱ��,Ϊ��������Ա����ṩ����,Ҳ��Ϊ����“ʮ�����”Ч�������ṩ�������ϡ�
1 �����뷽��
1.1 ������Դ
2016-2018�������������Դ��Ұ�����,��Ҫ���ڷ�����Ұӿ�����������ԵĿռ��֡�����վ�������Ůƺ(����)���罭(����)����Ϫ(��β)3������,�ֱ����ҰӴ��Լ11��90��123 km(ͼ1)����ˮǰ�����ζ�Ϊ��Ȼ��ˮ����,��ˮ�������Ůƺ���α�Ϊ��ˮ����;�����罭����Ϊ��ˮ�䶯��,ƽ������ԼΪ0.2��0.5 m/s,�ڿ�����ˮλ����ʱΪ��ˮ����,��ˮλ����ʱΪ��ˮ����;��β��Ϫ����Ϊ��ˮ����,ƽ�����ٴ���0.5 m/s[24]���������ʱ��Ϊ2016��7��10��,2017��4��8��11��,2018��8��10��,Ϊ�����ۺϱȽ�����Ⱥ��Ŀռ�仯,����ͬ����ʱ�ε����������һ����з���������������߰������ϴ����͵��������ϴ�����4����ͬ��Ŀ����Ƭ(��50 m,��2 m)����ƴ�Ӷ���,��Ŀ2a�ֱ�Ϊ2.0��6.0��10.0��14.0 cm��������18 m,��0.33 m,��0.45 m,��Ŀ2a=0.8 cm,ÿ��ʹ��3��������ÿ��վ��ÿ����������7��,ÿ����ҵʱ��Ϊ12 h(6:00 pm-6:00 am)��������,������״̬�¶����������������������ͳ��ء����������������ԭ�ط���,�����ĸ������������������������Ҫ�ο����Ĵ�����־��[25]��
ͼ1 ��Ұӿ������齭��Fig.1 Sampling sections in the Xiangjiaba Reservoir
��Ұӿ����罭����2011��(2011SJ)��2015��(2015SJ)������������Դ������[23],���2016-2018��ĵ�������(��Ůƺ2018SNP���罭2018SJ����Ϫ2018HX),������ˮǰ������Ұӿ�����������Եı仯���ơ�����,2011��Ϊ��ˮǰ�����,2015��Ϊ��ˮ���ڵ������
1.2 Ⱥ�������Է���
����Bray-Curtis (BC)���빹�������������ľ������,ͨ���Ƕ�����ά�߶�(non-metric multi-dimensional scale, NMDS)��������Ⱥ��ṹ��ʱ�ղ���,��в��ϵ��(stress)����NMDSͼ������,stress<0.2��ʾ��������һ����������,stress<0.1��ʾ�������Ϻ�,stress<0.05��ʾ������кܺõĴ�����[26-27]��
1.3 �����Բ�ȷ���
1.3.1 ���ֶ����� ����Margalef�ḻ��ָ��(D)��Shannon-Wiener������ָ��(H)��Pielou���ȶ�ָ��(E)������Ұӿ�������Ⱥ������ֶ�����[28],���㹫ʽΪ:
D=(S-1)/lnN
(1)
H=-∑WilnWi
(2)
E=H/lnS
(3)
ʽ��,S������������,N���ܸ�����,Wi�ǵ�i������ĸ�����ռ�ܸ������ı�����
1.3.2 ��������� ��������Կ����������ڷ���ѧ�ϵĹ�������,���ֹ����ڶ������Ⱥ������ֹ�����ͬһ����Ⱥ���и��ߵĶ����ԡ����÷��������ָ��(Δ, taxonomic diversity)�ͷ������ָ��(Δ*, taxonomic distinctness)����������Ⱥ��ķ��������[4]��������ָ���ȿ��������ֵ�����,Ҳ���������ֵķ��,���㹫ʽΪ:
(4)
(5)
ʽ��,Wij��ʾ����i��j�ڷ���ϵͳ��״ͼ�е�·������,Xi��Xj�ֱ��ʾ����i��j�ķ�ȡ�����Ŀ���ơ�������4������ȼ�ˮƽȷ������ķ����λ,��ͬˮƽ���������Ȩ��ֵΪ:Ŀ��Worder=100,�Ƽ�Wfamily=75,����Wgenus=50,�ּ�Wspecies=25[29]��
1.3.3 ���ܶ����� ͨ���������������������ݿ�Fishbase(https://fishbase.mnhn.fr/search.php)�ռ�������ʳ(food acquisition)���˶�(locomotion)�ͷ�ֳ(reproduction)����صĹ�����״,������λ�����͡���ֳ����(������)[30]���ο�������[31]���о���������̬���ͷ�Ϊ3��:(1)��ˮ��(limnophilic, L),��ֳ������������ξ��ھ�ˮ�н���;(2)��ˮ��(rheophilic, R),��Ϣ����ˮ������;(3)������(eurytopic,E),��ˮ��ˮ�ж��������档�����ռ����������������Ŀ�����ֵ���״��Ϣʱ,�ο���������ͬ������������ֵ���״[32]��
���ù��ܷḻ��(functional richness, FRic)�����ܾ��ȶ�(functional evenness, FEve)������ɢ��(functional divergence, FDiv)3��ָ������������Ⱥ�书�ܶ�����[30]�����ܷḻ��ָ����ʾijȺ��������ռ�ݹ��ܿռ�Ķ���,���ܾ��ȶȺ������������ڹ��ܿռ��еľ��ȷֲ����,��������ɢ���Ǻ������������仯��ָ��[30]�����ܷḻ�ȼ��㹫ʽΪ:
FRic=SFic/Rc
(6)
ʽ��,SFicָȺ����������ռ�ݵ���̬λ,Rcָ����ֵ�ľ���ֵ��
���ܾ��ȶȼ��㹫ʽΪ:
(7)
(8)
ʽ��,PEWiΪ��֧��Ȩ��,SΪȺ���е���������,dist(i,j)Ϊ���ֺ͵�ŷʽ����,lΪ��֧����
������ɢ�ȼ��㹫ʽΪ:
(9)
(10)
(11)
(12)
(13)
(14)
ʽ��,SΪȺ���е���������,xikΪ����i��״kֵ,gkΪ��״k������,TΪ��״��,Δ|d|Ϊ����i�����ĵ�ƽ������, ΔdΪ����Է��ΪȨ�ص���ɢ��,WiΪ����i����Է�ȡ�����,����Ⱥ��������Ȩƽ����ָ��(community-weighted mean, CWM)��Ϊ���ܶ����ԵIJ���[33],���㹫ʽΪ:
(15)
ʽ��,traitsiΪ����i����״ֵ,SΪȺ���������������ڸ�Ⱥ�����ֵ���Է�Ⱥ�����״ֵ�ֱ������ʳ���˶��ͷ�ֳ���ܵ�CWMֵ��
1.4 ���ݷ���
ʹ��EXCEL�����������������ֶ�����ָ������,ʹ��PRIMER 5.0����������������ָ����NMDS�����ܶ�����ָ���Ͷ�����ָ���������Էֱ����R 4.1.0�����е�“vegan”��“FD”��“Hmisc”���������з�����
2 ���
2.1 ����Ⱥ�����
2016-2018������Ұӿ���3�����ι��ɼ�����59��5016β,������6Ŀ16��48��������Ůƺ���罭����Ϫ���ηֱ�ɼ�������39��40��39�֡���Ŀ������ˮƽ����,����Ŀ�������,��36��,ռ��������61.0%;���������Ŀ,��13��,ռ��������22.%;����Ŀ��7��,ռ��������11.8%,����Ŀ������Ŀ��������Ŀ��1��,�ֱ�ռ��������1.6%(������)������,������������������5��,����������12�֡���Ůƺ������(Hemiculterleucisculus)����������(Rhodeusocellatus)���л�������(Spinibarbussinensis)Ϊ��,�����ٷֱȷֱ�Ϊ27.95%��16.22%��12.53%;�罭�����Ժ����ϻ����(Pelteobagrusvachelli)Ϊ��,�����ٷֱȷֱ�Ϊ45.94%��12.45%;��Ϫ���������ϻ�����Ϊ��,�����ٷֱȷֱ�Ϊ25.73%��16.55%(��1)��������,��Ұӿ��������Թ����Ե����ϻ����;�ˮ�Եĵ�ռ����,���ߵ������ٷֱȷֱ�Ϊ30.15%±14.82%��13.91%±11.16%��Բ��ͭ��(Coreiusguichenoti)���л�ɳ��(Botiasuperciliaris)����ˮ�������ڿ�β��Ϫ�����зֲ�,���ڿ�����Ůƺ�Ϳ����罭δ�ɼ�����
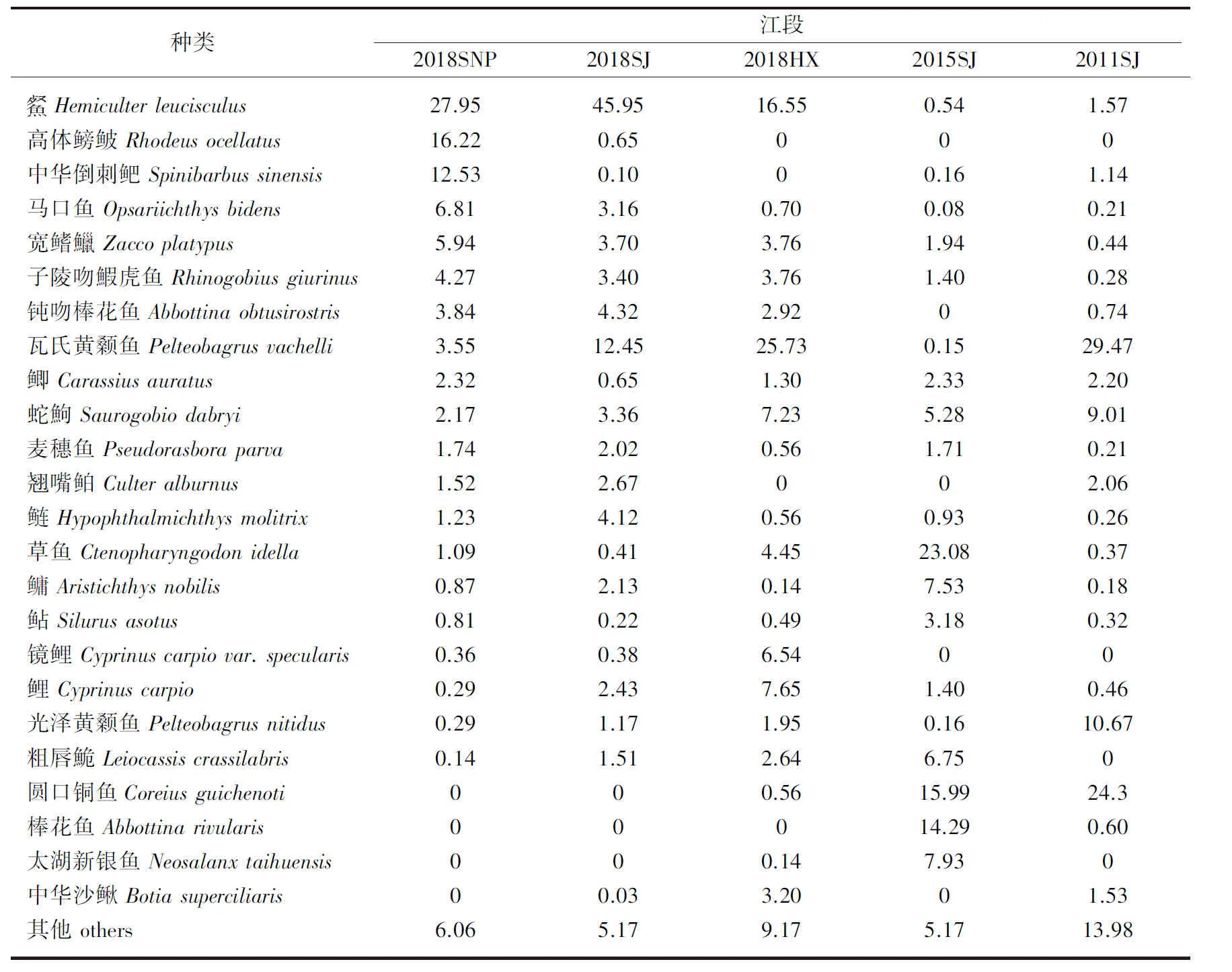
��1 ��ˮǰ����Ұӿ�������Ⱥ�������ٷֱ����(%)Tab.1 Composition of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
��ˮǰ����ˮ���ں�2016-2018��ֱ����罭���βɼ�������54��35��40�֡���ˮǰ�����ϻ���㡢Բ��ͭ���������(P.nitidus)ռ����,�����ٷֱȷֱ�Ϊ29.47%��24.30%��10.67%����ˮ�����Բ���(Ctenopharyngodonidella)��Բ��ͭ��Ͱ�����(Abbottinarivularis)ռ����,�����ٷֱȷֱ�Ϊ23.08%��15.99%��14.29%��2016-2018���Ժ����ϻ����ռ����,�����ٷֱȷֱ�Ϊ45.95%��12.45%(��1)����Ұ���ˮ��,��ɳ��������Ұӿ����������ı仯��Բ��ͭ�����ˮ�������½�,�ڿ��Ϳ�����ʧ,ֻ�ڿ�β��ˮ���λ��зֲ�,���Ⱦ�ˮ����������ˮ���ڱ�Ϊ�����֡�
��ͼ2���Կ���,�ռ���,��ˮ����Ұӿ���������Ůƺ�Ϳ����罭����Ⱥ�������С,������Ůƺ�Ϳ�β��Ϫ����Ⱥ��������ʱ����,�����罭����ˮǰ����ˮ���ں�2016-2018��3��ʱ�ڵ�����Ⱥ�������ϴ�,������ˮ���罭��������Ⱥ�侭���˾��ұ仯����ˮ���β��Ϫ����ˮǰ�罭����Ⱥ��֮�����������Խϸߡ�
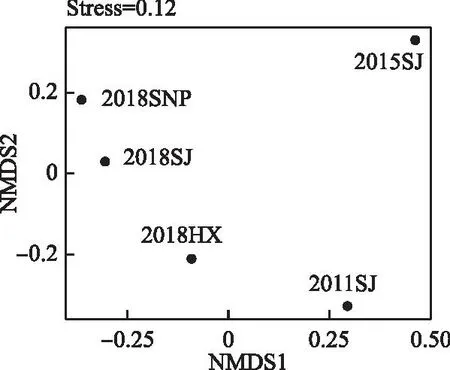
ͼ2 ��ˮǰ����Ұӿ�������Ⱥ��NMDSͼFig.2 Non-metric multi-dimensional scale ranking of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
2.2 ���ֶ�����
2016-2018����Ұӿ�������Ⱥ��Margalef�ḻ��ָ���ı仯��ΧΪ4.89��5.78,ƽ��ֵΪ5.31±0.45;Shannon-Wiener������ָ���ı仯��ΧΪ2.20��2.64,ƽ��ֵΪ2.45±0.23;Pielou���ȶ�ָ���ı仯��ΧΪ0.60��0.73,ƽ��ֵΪ0.67±0.06���ռ���,�������ֶ�����ָ��������Ϊ:2018HX>2018SNP>2018SJ,������β��Ϫ�����������ֶ��������,�����罭�������(ͼ3)��
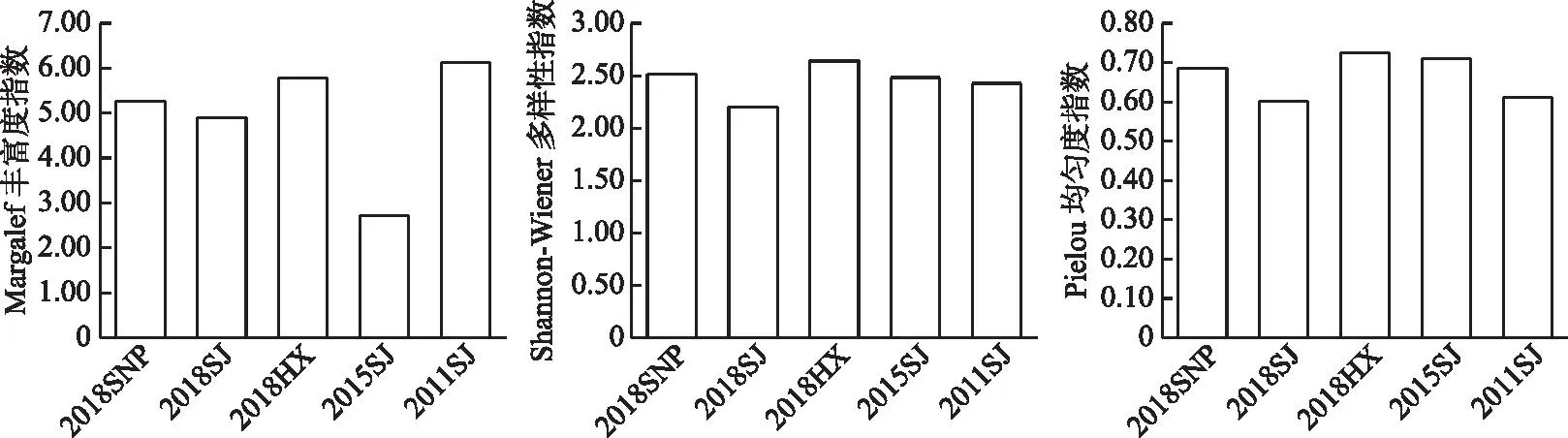
ͼ3 ��ˮǰ����Ұӿ�������Ⱥ�����ֶ����Ա仯Fig.3 Changes of species diversity of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
��ˮǰ��Աȱ���,�����罭����Margalef�ḻ��ָ����ˮǰ���(6.13),��ˮ�����½�����(2.72),����Ϊ4.89��Shannon-Wiener������ָ����ˮ���ڱ仯����,�����½���Pielou���ȶ�ָ����ˮ��������,���ڻָ�����ˮǰˮƽ��������,��ˮ���罭�����������ֶ����Ա���Ϊ�½����ơ�
2.3 ���������
2016-2018����Ұӿ�������Ⱥ����������ָ���ı仯��ΧΪ53.39��70.77,ƽ��ֵΪ59.56±9.72,�������ָ���ı仯��ΧΪ61.70��80.15,ƽ��ֵΪ71.13±9.23����β��Ϫ���ε�Ŀ���ơ����������Ը��ڿ�����Ůƺ�Ϳ����罭����(��2),��������������ָ�����������ָ�������,�ֱ�Ϊ70.77��80.15(ͼ4)���ռ���,������������ָ�����������ָ���ı仯����һ��:2018HX>2018SJ>2018SNP,������β��Ϫ�������������������,������Ůƺ������͡�
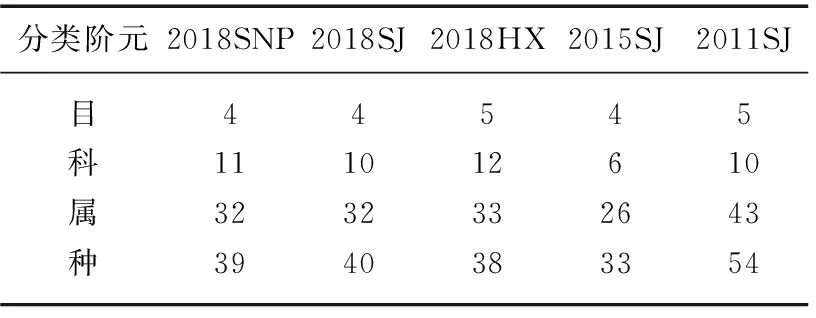
��2 ��ˮǰ����Ұӿ�������ķ������Tab.2 Fish taxonomic composition in the Xiangjiaba Reservoir before and after impoundment
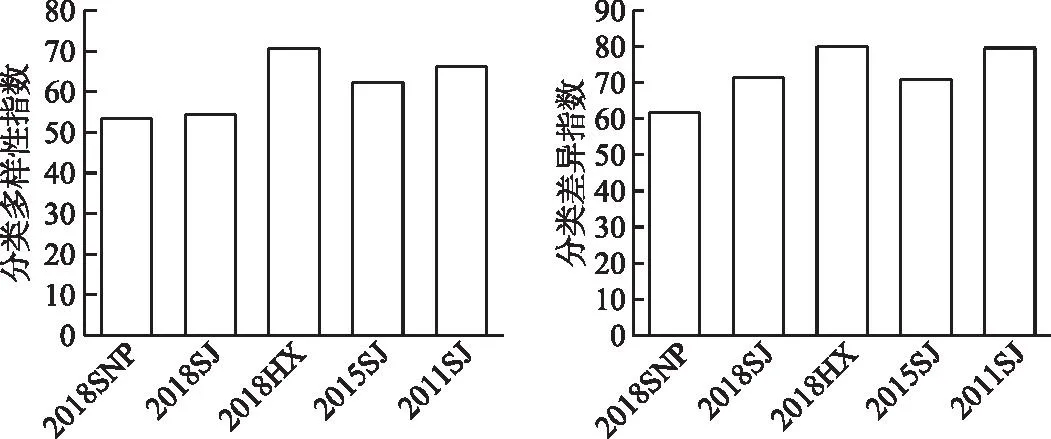
ͼ4 ��ˮǰ����Ұӿ��������������Ա仯Fig.4 Changes of taxonomic diversity of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
��ˮǰ��Աȱ���,�����罭��������IJ�ͬ�����Ԫ����������Ϊ��ˮ�����½����������������ơ���ˮ���罭���η�������Ա���Ϊ�½�����,���������ָ������ˮǰ��66.16�½�Ϊ��ˮ���ڵ�62.29,2016-2018���½�Ϊ54.53;�������ָ������ˮǰ��79.63�½�Ϊ��ˮ���ڵ�70.92��
2.4 ���ܶ�����
2.4.1 ��ʳ���ܶ����� 2016-2018����Ұӿ���������ʳ���ܷḻ�ȱ仯��ΧΪ0.010��0.011,ƽ��ֵΪ0.011±0.000;��ʳ���ܾ��ȶȱ仯��ΧΪ0.46��0.56,ƽ��ֵΪ0.50±0.05;��ʳ������ɢ�ȱ仯��ΧΪ0.81��0.90,ƽ��ֵΪ0.85±0.05(ͼ5)���ռ���,3����������Ůƺ������ʳ���ܷḻ�����,����ʳ���ܾ��ȶ����,������������ʳ������ɢ����������ʳ������״����,3�����ξ�Ϊͷ��/�dz���ͷ��/�峤��ͷ��/�ۼ��ߵ��������ʳ������ռ����;��Ůƺ���罭���Զ�λ�ڵ�����ռ����,��Ϫ����λ�ڵ�����ռ����;��Ůƺ����Ϫ�������־�Ϊ������ˮ�²������,�罭���ε�������Ϊ������ˮ�ϲ������;�ӿ�����Ůƺ����β��Ϫ����,�����ֵ�Ӫ������ͷ��/�۾�����Ϊ�����ӵ����ơ�
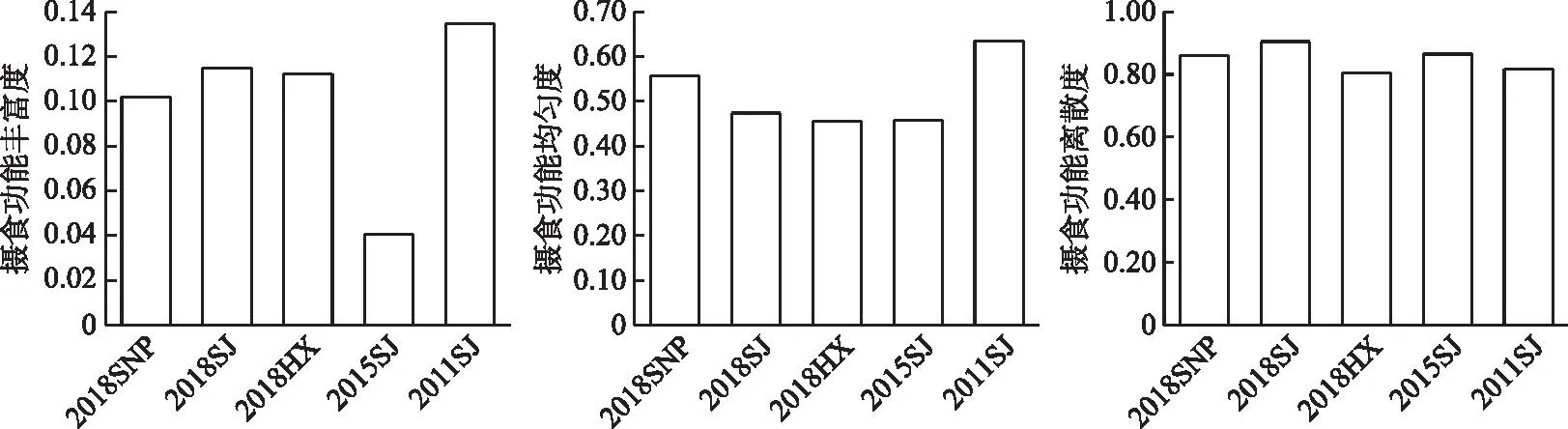
ͼ5 ��ˮǰ����Ұӿ���������ʳ���ܶ����Ա仯Fig.5 Changes of food acquisition functional diversity of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
����ˮǰ���,��ˮ���罭����������ʳ���ܵķḻ�Ⱥ;��ȶȾ��½�,�ֱ���0.013��0.63�½���0.011��0.47,��ʳ������ɢ�ȱ仯��������ʳ������״������Ҫ����Ϊ:�����ֵ�Ӫ������ͷ��/�۾����½�����,�ֱ���3.39��6.80�½���2.98��4.32;��λ��������ˮǰ����λ��ռ���Ʊ�Ϊ��ˮ��Ķ�λ��ռ����,����ˮ�����²�ռ���Ʊ�Ϊ�ϲ�ռ����(��ͼ��)��
2.4.2 �˶����ܶ����� 2016-2018����Ұӿ��������˶����ܷḻ�ȵı仯��ΧΪ0.007��0.012,ƽ��ֵΪ0.009±0.002;�˶����ܾ��ȶȵı仯��ΧΪ0.35��0.49,ƽ��ֵΪ0.43±0.07;�˶�������ɢ�ȵı仯��ΧΪ0.87��0.94,ƽ��ֵΪ0.91±0.04(ͼ6)���ռ���,3���������罭���������˶����ܷḻ�����,���˶����ȶ���͡����˶�������״����,3�����ξ����峤/��߱�ֵ�ͺͲ��������ռ����;��Ůƺ����Ϫ��Ϊ����������ռ����,�罭�����Ծ�ˮ������ռ����;�����ֵ�����峤���峤/β�������峤/β���ߴӿ�����Ůƺ����β��Ϫ���������ơ�
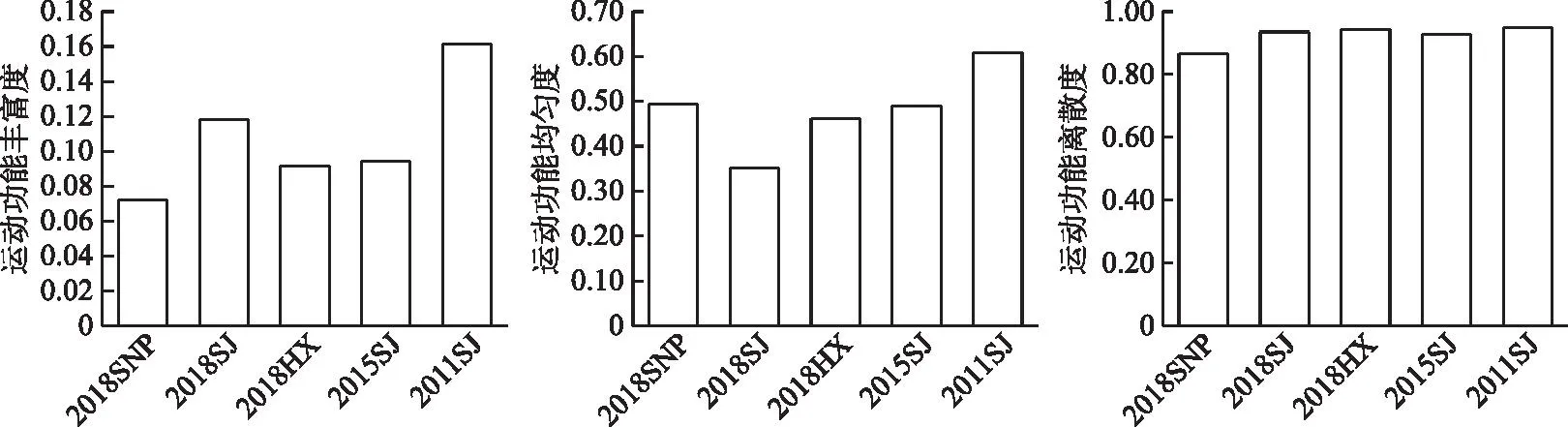
ͼ6 ��ˮǰ����Ұӿ��������˶����ܶ����Ա仯Fig.6 Changes of locomotion functional diversity of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
����ˮǰ���,��ˮ���罭���������˶����ܷḻ�Ⱥ;��ȶ��½�,�ֱ���0.16��0.61�½���0.12��0.35,�˶�������ɢ�ȱ仯�������˶�������״������Ҫ����Ϊ:�����ֵ�����峤�½�����,��32.32 cm�½���24.37 cm;������������ˮǰ�Ĵ���ռ���Ʊ�Ϊ��ˮ���ڵ�ԲͲ��ռ����,2016-2018���Բ����ռ����;������̬�����ɹ�����ռ����ת��Ϊ��ˮ��ռ����(��ͼ��)��
2.4.3 ��ֳ���ܶ����� 2016-2018����Ұӿ�������Ⱥ�䷱ֳ���ܷḻ�ȵı仯��ΧΪ0.124��0.239,ƽ��ֵΪ0.170±0.060;��ֳ���ܾ��ȶȵı仯��ΧΪ0.36��0.54,ƽ��ֵΪ0.46±0.09;��ֳ������ɢ�ȵı仯��ΧΪ0.76��0.77,ƽ��ֵΪ0.76±0.003(ͼ7)���ռ���,3���������罭���ֳ���ܷḻ�����,����ֳ���ȶ���͡��ڷ�ֳ������״����,3�����������ֶ�Ϊ�����Գ������䶼��С����ճ���Ѻ��ļ����ѵ�����;�罭���ε�ƽ����Է�ֳ������������������;�����ֵij����Գ����峤��ƽ�����Է�ֳ���ӿ�����Ůƺ����β��Ϫ���γ��������ơ�
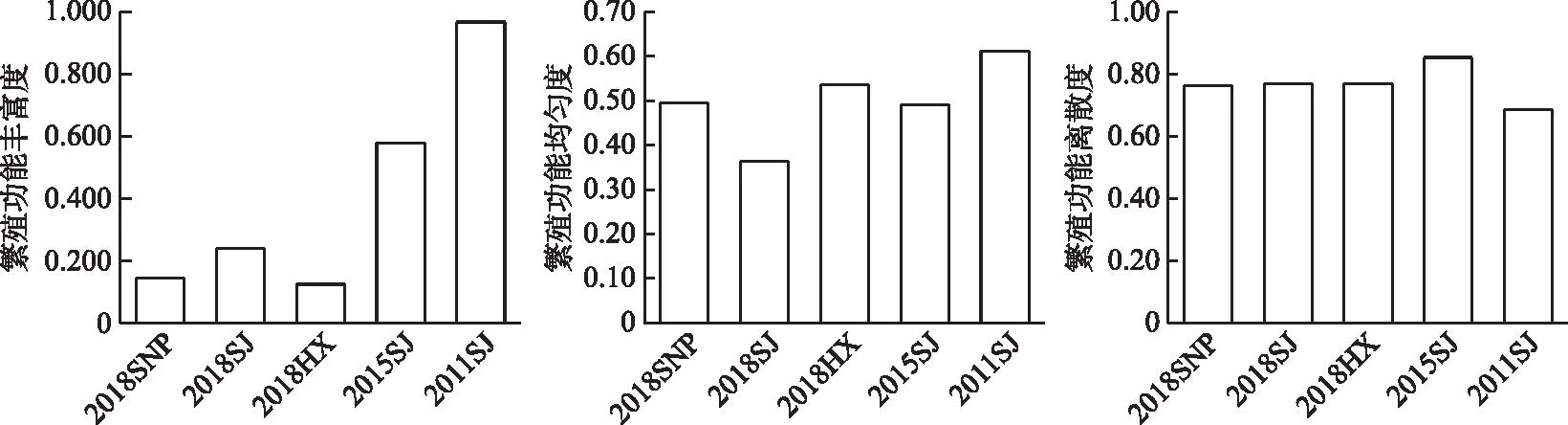
ͼ7 ��ˮǰ����Ұӿ������ֳ���ܶ����Ա仯Fig.7 Changes of reproduction functional diversity of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
����ˮǰ���,��ˮ���罭�������ֳ���ܷḻ�Ⱥ;��ȶȳ������½�����,�ֱ���0.97��0.61�½���0.24��0.36;��ֳ������ɢ�ȳ���������,��0.69������0.77���ڷ�ֳ������״������Ҫ����Ϊ:�����Գ����峤�ͳ����Գ��������½�,�ֱ���19.35 cm��2.31���½���14.36 cm��1.41��;ƽ����Է�ֳ����ƽ�����Է�ֳ������,�ֱ���393��/g��23525��������5722��/g��49250��(��ͼ��)��
2.4.4 �ܹ��ܶ����� 2016-2018����Ұӿ�������Ⱥ���ܹ��ܷḻ�ȵı仯��ΧΪ0.004��0.005,ƽ��ֵΪ0.004±0.000;�ܹ��ܾ��ȶȵı仯��ΧΪ0.43��0.50,ƽ��ֵΪ0.47±0.04;�ܹ�����ɢ�ȵı仯��ΧΪ0.86��0.93,ƽ��ֵΪ0.91±0.04(ͼ8)���ռ���,3���������罭�ܹ��ܷḻ�����,���ܹ��ܾ��ȶ���͡�
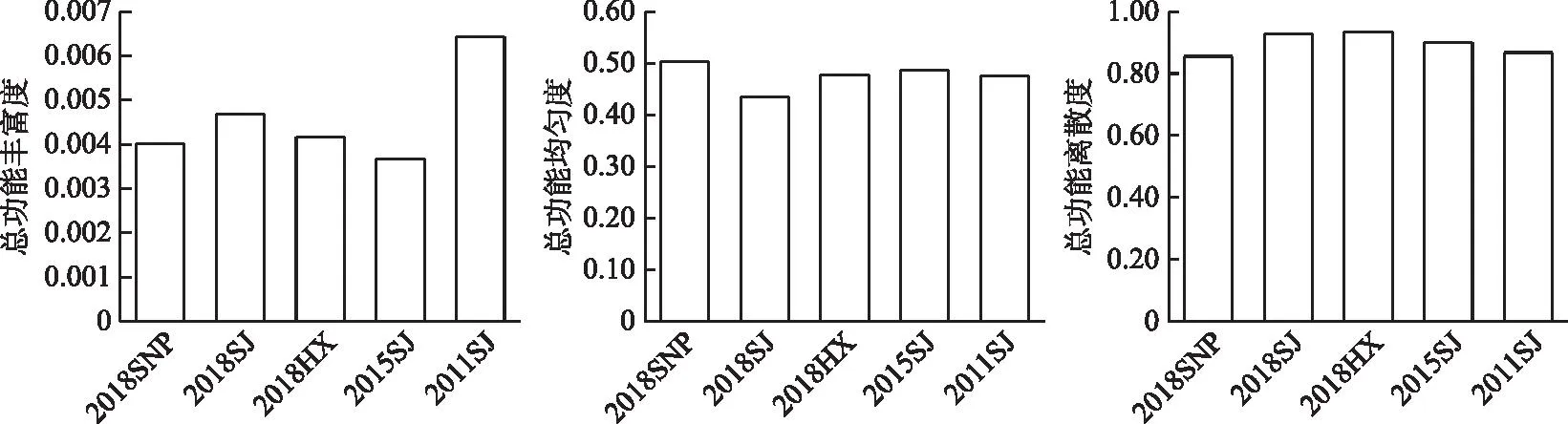
ͼ8 ��ˮǰ����Ұӿ��������ܹ��ܶ����Եı仯Fig.8 Changes of total functional diversity of fish assemblages in the Xiangjiaba Reservoir before and after impoundment
����ˮǰ���,��ˮ���罭���������ܹ��ܷḻ�Ⱥ;��ȶȳ��½�����,�ֱ���0.006��0.48�½���0.005��0.43,�ܹ�����ɢ�ȱ仯����
2.5 ������ָ�������ط���
Pearson��ط�����ʾ,��Ұӿ�����ͬ��������Margalef�ḻ��ָ������ʳ������ɢ�ȳ����������(n=3,r=0.97,P<0.05),��ֳ���ܾ��ȶ���Shannon-Wiener������ָ����Pielou���ȶȳ����������(n=3,r=-0.97,P<0.05;n=3,r=-0.97,P<0.05),�������֡����ࡢ���ܶ�����ָ��֮����ع�ϵ������(P<0.05)����ˮǰ���罭��������Shannon-Wiener������ָ�����ܹ��ܾ��ȶȳʼ����������(n=3,r=0.99,P<0.01),�������ָ������ʳ���ܾ��ȶȳʼ����������(n=3,r=0.98,P<0.01),���˶�������ɢ�ȳ����������(n=3,r=0.92,P<0.05)��������,��Ұӿ����������֡�������ܶ����Ը�ָ��֮����ع�ϵ��������
3 ����
3.1 ��������Կռ���
��ɳ��������Ұӵ�վ���ɺ�,ԭ����Ͽ�Ⱥ�������������ɺ���Ρ����ɶκͺ������ɵĸ�������[24,34],������Ůƺ�Ϳ����罭�������ھ�ˮ����,����β��Ϫ����Ϊ��ˮ����[35]����ˮ������ɥʧ,��������ˮ��������Ⱥ�����½�������ʧ[36]������������ˮ��������������ת��,�Ⱦ�ˮ���������������������������������ӡ�һ����Ϊ,��Ϣ�ػ����ı�ᵼ�²���Ӧ��������ʧ,�����������ַḻ��[37-38]����Ұӿ�����Ůƺ�������罭�Ϳ�β��Ϫ���ε���������������,����Ҫ����Ϊ��ˮ��������Ⱥ�������ȶ�,����������ˮ��������Ȼ����,���Ǿ�ˮ�Ժ�������������[39-40]�����о�����,�ڿռ�ֲ��Ͽ�β��Ϫ���ε����ֶ��������,�������õȵ��о����һ��[24]����ˮ��,��β���������Ծ���һ����Χ����ˮ����,���нϸߵ�����������,�ܹ�֧�Ÿ��ߵ����������[35,41-42]�������ڼ�,��ˮ�Ե�Բ��ͭ����л�ɳ�����ڿ��Ϳ��н��ξ�����ʧ,���ڿ�β��Ϫ������Ȼ��һ������������,���о����Ҳ��ʾ,��β��Ϫ���ε�������������Ҳ���ڿ��кͿ����Ρ���ٻ�õ�[43]�о�����,��ˮ������Ϣ�������Խ���,�������ͬ�ʻ�,�����ε������������Խ������ԡ�
���о����ֿ����罭���ε�������ʳ���˶��ͷ�ֳ��ع��ܷḻ�Ƚϸ�,��������罭������������,���ֽ�Ϊ�����йء��罭Ϊ��Ұӿ����Ĺ��ɶ�,�ڿ�����ͬˮλ����ʱ����ڻ���ˮ�;�ˮ��������[24]�����ӵ���������ӵ�в�ͬ��״�������,�����ܷḻ��ֻ���������ֱ����Ĺ�����״,����Ȩ�����ֶ��[44-45]�����,�罭���ε����ֹ�����״�ķ�Χ����,���ܷḻ�ȸ��ߡ��빦�ܷḻ�Ȳ�ͬ,�罭���ε�������ʳ���˶��ͷ�ֳ��ع��ܾ��ȶȽϵ�,���������Ϊ���ܾ��ȶȺ���������״�ڹ��ܿռ�ֲ��ľ��ȳ̶�,�����ڹ�����״ֵ�����ֵ���Զ��[45]���罭���������ֺ����ϻ����������ٷֱȷֱ�ߴ�45.94%��12.45%,������������ռ�ȶ�����5%,��Է�Ȳ�����ᵼ�������ڹ��ܿռ��Ϸֲ������ȡ���Ұӿ�������������Ĺ�����ɢ��������ֵ�Ƚϸ�,˵����ˮ���������Ⱥ�������ȶ�״̬,���ּ侺������,��̬λ�ֻ��̶Ƚϸ�,��ͬ�����ֶ������Բ�ͬ�ķ�ʽ����Ⱥ���е���Դ[46],��Ҳ�Ӳ���˵��Ⱥ����ֻ���ٲ��ֵ���Դ����Ч���ö������ϵ���Դ���ò����,���ܾ��ȶȽϵ͡�
3.2 ��ˮ����������Ա仯����
��ˮ����Ұӿ������ֶ����Ա���Ϊ�½�����,������ˮ��������ε�ˮ�Ļ����仯�йء���ˮ����ˮ������Ͳ�Ư�����ѵ������������˻����ĸı������ʧ��������ˮ����Ǩ��,�������ֶ������½�[47]��Ф����[40]�о����ڽ��غ�����������������ˮ��վ��ˮǰ����ˮ��������ɼ����ֶ����Եı仯,������ˮ����齭�ε����ֶ����������½�,��������100���½�����62��,Margalef�ḻ��ָ����Shannon-Wienerָ��Ҳ�ֱ��½���43.35%��60.80%��ˮ��վ�Ľ�������������ı�����黯,��������ԭ�е���ˮ�������ģ����,����û�˲�������IJ��ѳ�,�������ֶ����Է����仯[40]�����о���ʾ,��ˮ����Ұӿ�����������Ⱥ��ķ��������Ҳ���ֳ��½����ơ�������Ⱥ���ܵ���Ϊ����ʱ,Ⱥ�������ּ�Ľ�����ϵ�ᷢ���ı�[4]��Clarke����Ϊ[48],�������ܵ�����ʱ,����Ⱥ���а������������ٵ������Ƶȸ����Ԫͨ����������ʧ,����Щ�������ַḻ�������Ƶ��������ܹ�����,��ʹȺ��ķ�������Խ���,����������½���
��ˮǰ�罭����������ʳ���˶��ͷ�ֳ���ܶ����Զ��ǽϸߵ�,����ܺ���ˮǰ�ý���������Դ�ḻ,Ⱥ���ȶ��йء����ܶ�������ָ�ض���̬ϵͳ�����ֹ�����������ֵ�ͷ�Χ,ǿ������Ⱥ�������ֹ��ܵIJ���[5,44],��Ⱥ����û���κ��������־�����ȫ��ͬ�Ĺ�������,������������Խ��,Ⱥ���Ĺ��ܲ���Ҳ��Խ��ͬʱ,���ڹ�����״����̬����Ӱ������,�����ĸı�Ҳ��Ӱ�칦�ܶ����ԡ���ˮǰ��Ұӽ���Ϊ��Ȼ��ˮ����,Ⱥ��������Ӧ��ˮ������Ϊ��,��Բ��ͭ�������Ϊ�Ĵ��ε�����,���н�ǿ����Ӿ��������ˮ���������ټ���,����Ӿ������Խ����IJ����������Ϊ����������[49]��������ˮ��,�ı���ԭ����������,��������������ɷ����ı�,ʹ����ܶ����Խ��͡�Oliveira��[50]�о��˰���Paraná������Ⱥ�������δ�ӽ���ǰ�ͽ������ܶ����Եij��ڱ仯,���ִ�ӽ��������κӶ������ı�,��Ⱥ����������ɲ�������Ӱ��,��������Ⱥ�书�ܼ�,���ܶ����Խ��͡�Zhang��[51]�о���Ͽ�����ˮǰ����ˮ�ڼ����ˮ������������Ⱥ�����ֶ����Ժ��ܶ����Եı仯ʱ����,��ˮ��Ⱥ���ڲ��ȶ���״̬��������������������,����Ⱥ��ṹ�����仯,ʹ���ܶ����Խ��͡�
3.3 ���֡�������ܶ�����֮��Ĺ�ϵ
�о�����,���ֺͷ��������֮��Ĺ�ϵ���ܻ�����Ӱ������ֳ���ͬ�Ľ��[52]��Heino��[53]�о��˷�����ͬ�����ͺ������������ֺͷ�������ԵĹ�ϵ,���ֲ�ͬ���������ߵĹ�ϵ����ܴ�,����������ء���������غͲ���ص�,�ڲ�ͬ������Ӱ�����߹�ϵ�����Ӳ�ͬ��ͬ��,���ֺ��ܶ����ԵĹ�ϵ�ڲ�ͬ��Ⱥ��ͻ�����Ҳ���ܴ��ڲ�ͬ�Ľ��,��������ء���������غͲ���ص�[54-56]��Mayfield��[57]��Ϊ,��Ⱥ���й��������ķ�Χ�������ֵ����Ӷ�����ʱ,���ַḻ�Ⱥ��ܶ�����Ϊ�����;���������ֶ����Ժ��ܶ����ԶԻ����ݶȵ���Ӧ�̶Ȳ�ͬ,����֮��Ĺ�ϵҲ�ᷢ���ı�[58]��Cadotte��[59]�ع���Ӱ�����ֶ����Ժ��ܶ����Թ�ϵ�ķ���������,��Ӫ���Ρ��¶Ⱥ��Ŷ���,�����ڲ�ͬ����Ӱ�������ֺ��ܶ����ԵĹ�ϵ����Ϊ������غͲ���ء�����,��������Ҳ��Ӱ�칦�ܶ����Ա仯����Ҫ����,�ڹ�������ߵ�Ⱥ����,��ʹ���ֶ����Խ���,���ܶ�����Ҳ�����������͡����о������֡�������ܶ����ԵĶ�����ط�����ʾ,���ߵ�����Բ�ǿ,������������ƫ���й�,Ҳ�����벻ͬά�ȶ����ԵIJ��ص㲻ͬ�й�,���ֶ�����ǿ��Ⱥ���ڲ������������������������Ϣ,��������Բ��������ּ�ķ����ϵ����Ȩ��,���ܶ���������ǿ��������״����̬ϵͳ�Ĺ���[60]��
���о�������ˮ������Ұӿ���������ɷ������Ըı�,�������֡�������ܶ����Զ����ֳ��½������ơ����,�ڳ���“ʮ�����”�ı�����,Ӧ��ǿ��ɳ��������������Ա�������̬����һ����,��ǿ����Ұӿ�β��ˮ���ε�������������,Ϊ��ˮ�������ṩ���˵���������;��һ����,���鿪չ�ݼ���վ��������̬����,ά����������ͷ�ֳ�������̬���������֡�������ܶ����Էֱ�ӳ����������ԵIJ�ͬ����,�ڽ�������Ⱥ���о���,��϶���ָ�����������ڸ�ȫ����˽�����Ⱥ��Ի����仯��������ŵ���̬��Ӧ��
4 ��¼
�������ͼ������Ӱ�(DOI: 10.18307/2024.0132)��