
������ԣ��������� �Ų�ԭ��
������Ƭ��ע��ǧ��-������������ӳ�Ա-�й���������ѧ���о��ң�
�ڹ���ǭ��������ˮ�������ض�����ˮ�����ش����Ұ������������
����“������Ų�ԭ��”��1�š���2�����Ϲ۲������
�岥������������Ƭ����������Ա���Ҷ������䡢����
ԭ���⣺����Ұ������������״��������ֶ����Է���
� ���� �Ǵ�� ���� ��˼�� ���� ������

ժҪ��Ϊ���õر����Ϳ�����������Ұ��������Դ����16������Ұ������Ϊ�о������ø�ЧҺ��ɫ������ֹ��ȷ�����������Ҫ������֣����ڴ������^�Ա�����״��չ�۲⣬��ϱ���ϵ�����Ŵ�������ָ����������Ұ��������Դ����̶ȺͶ�����ˮƽ�����������20�������Ա�����״����ϵ����0��35.68%֮�䣬ƽ��ֵΪ9.94%���Ŵ�������ָ����0��1.095֮�䣬ƽ��ֵΪ0.257��9�������Ա�����״����ϵ����0��32.60%֮�䣬ƽ��ֵΪ12.68%���Ŵ�������ָ����0��1.927֮�䣬ƽ��ֵΪ1.077��17��������״����ϵ�����Ŵ�������ָ������ֱ�Ϊ0��400.0%��0��1.927��ƽ��ֵ�ֱ�Ϊ65.28% ��1.320������Ұ��������Դ���ͼ�����״����Ϊ�ӷ�3�ҡ���ë����ѿ��ë��������������Ϊ�߲��ӡ���C���߿ɿɼ�͵����백���ᡢ�谱�ᡢ�Ͷ�������������EGCG���ͻ����ȼ����Ұ��������Դ������״��������ֽ�ͳһ����Դ�ڲ�����С��C�Ϳɿɼ�Ϊ����������֡���ϱ��ͺ������������Ʋ�����Ұ��������Դ����Ϊ�Ž���ı��֡�
�ؼ��ʣ�������Ұ���������Ŵ�������ָ�����ɿɼ������״���������
��ͼ����ţ�S571.101 ���ױ�־�룺A
���±�ţ�1002-1302��2023��08-0111-08
������Ŀ ����ʡ�Ƽ��ƻ�����ţ�ǭ�ƺ�֧�ţ�2019��2254�š�ǭ�ƺ�ƽ̨�˲ţ�2020��2103�ţ�������ʡũҵ��ѧԺ��ɫ��ֲ����Դ�ռ��������봴��������Ŀ����ţ�ǭũ��Ժ������Դ��2022��03�ţ���
����飺�� ����1988—����Ů�����������ˣ�˶ʿ�������о�Ա����Ҫ���²�����Դ������Ʒ�ֿ����о���E-mail��610681998@qq.com��
ͨ�����ߣ������䣬�о�Ա����Ҫ���²�����Դ���ֹ�����E-mail��zwchentea@163.com��
�ҹ��Dz�����Camellia sinensis ��L.�� O.Kuntze�ݵ�ԭ���أ����ϡ����ݵ�ʡ�ֲ����������ࡢ���Ͷ�����Ұ��������Դ��1-4�ݣ���ЩҰ��������Դ���ڲ������Ŵ��ݻ���Ʒ�ָ������������ֵ��о�������Ҫ�����5-7�ݡ���ˣ��Ӵ����ȱ����������о���ϡҰ��������Դ�����ֽ����������Դ�о����ص��8�ݡ�
����ˮ�������أ�λ�ڹ���ʡǭ�ϲ��������������ݣ��ش�����ɽ����ɽ���أ��ǹ���Ұ���������зֲ���֮һ��9�ݡ�ǰ�����������Ұ��������Ե��ϵ���Ŵ������Ե�չ����һ���о���10-11�ݣ�����δ������Ұ����������������������״�ȷ������ر��������о�ѡȡ16�ݴ���������Ұ������Ϊ�о�����ͨ���ⶨ��Ҫ�����ɷֺ۲���������Ҷ�ȱ�����״�����ͳ�Ʒ�����������������Ұ��������Դ���ͼ������ı���̶ȺͶ�����ˮƽ�����ɳ�����Ұ��������Դ����ı�����������������������Ϊ��һ�������Ϳ�����������Ұ��������Դ���»�����
1 �����뷽��
1.1 �������Լ�
��ЧҺ��ɫ�ף�HPLC���ǣ�Agilent 1260 Infinity �����������ף���
����ֹ��ȼƣ�TU-1800��������������
ɫ����������������Hypersil ODS2��4.6 mm×250 mm��5 μm����
���ȼûʳ���ᡢ��������۶����أ�C�����������أ�EC������ûʳ�Ӷ����أ�EGC������������ûʳ��������ECG����ûʳ�Ӷ�����ûʳ��������GCG������ûʳ�Ӷ�����ûʳ��������EGCG�� �ݺͲ谱���Ʒ���ԺϷʲ�������Ƽ�����˾��N��N-���������������桢�״�����������л��Լ�Ϊ����HPLC����
1.2 �������
�������ȡ�Թ���ʡǭ��������ˮ��������ԭʼɭ���У�������ΧΪ25°55′48″��25°59′40″N��108°9′32″��108°20′46″E�����η�ΧΪ1 300��1 500 m��ѡȡ��������10 cm��Ұ����������ȡ�����ƣ�ͼ1����������ײ裨���գ�ȡ�Թ���ʡ��Ҷ�о�����̶������Դ�ԡ�
1.3 ���鷽��
1.3.1 ��Ʒ�Ʊ� ��2020��5�£��ɸ���ǵ���1ѿ2Ҷ��Ҷ������ɱ��ķ�����12�ݣ��Ʊ����������������ȡ��ʱ��Ϊ2020��4�£�ȡ����1ѿ2Ҷ��
1.3.2 ��ⷽ��
������״���ֱ���2020�ꡢ2021�괺�����^�����¼����Ҷ������ѿ����״�����͵��鼰��������ο�������������Դ�����������ƹ淶�������û���ʿ������������VHX-5000���۲ⶥѿ���ӷ���Ҹѿ��ë��ÿ�ݲ��ϵ���10��Ҷ��ѿ�����������Ա�����״ȡƽ��ֵ�������Ա�����״�Գ��ָ���������������Ϊ��
�����ɷ֣����백���Ậ������GB/T 8314—2013 ���� ���백���������IJⶨ���еķ������вⶨ�����Ӻ�������GB/T 8313—2018 ����Ҷ�в��ӺͶ�������ļ�ⷽ�������вⶨ���谱�Ậ���ο���漵ȵķ�����13�ݽ��вⶨ����������ֲο���˳��ȵķ�����14�ݽ��вⶨ��ÿ����Ʒ�ظ����3�Σ�����ȡ3��ƽ��ֵ��
1.3.3 ���ݴ���
�����Ŵ�������ָ��������ũ-ά��ָ����Shannon-Wiener index������״�ֲ�Ƶ�ʼ������Է�������Excel 2010���������������Pearson����Է�������SPSS 21.0��
2 ��������
2.1 ������״�Ŵ������Է���
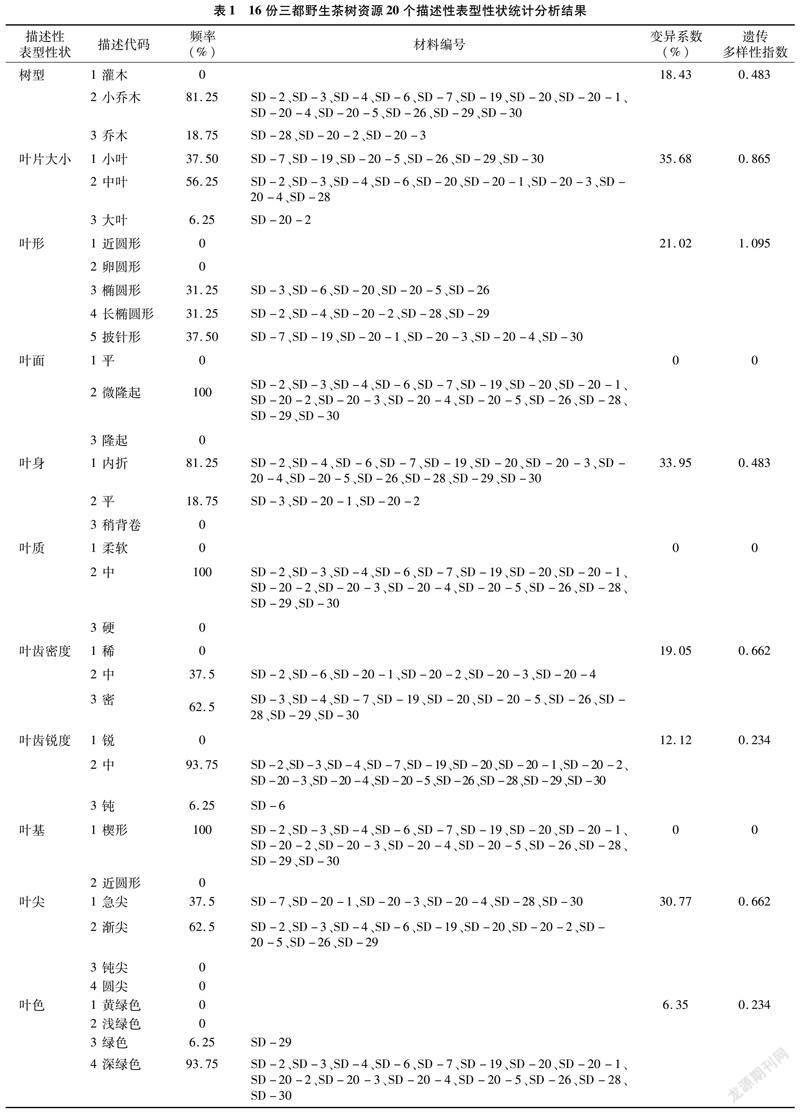
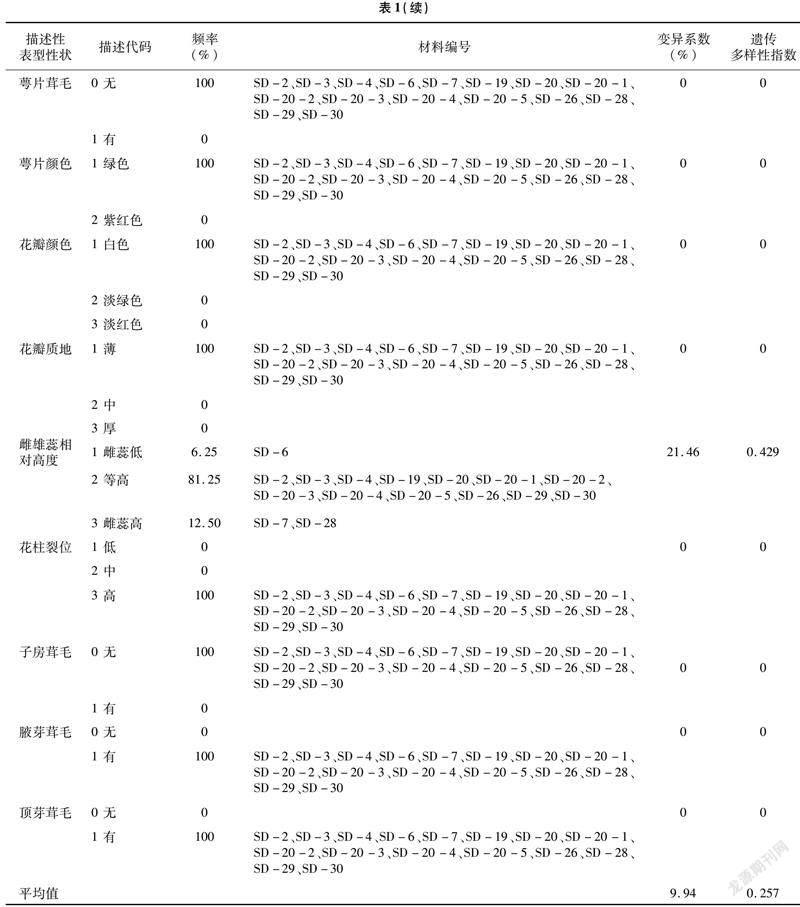
���о���ͳ�Ʒ�����16������Ұ���������͡�ҶƬ��С��Ҷ�ε�20�������Ա�����״��Ҷ����Ҷ����9�������Ա�����״����1����2����20�������Ա�����״�������������в�ͬ�ķֲ����֣�����ҶƬ��С�ʹ�������Ը߶������������Ͼ��зֲ��������ȣ�����ҶƬ��С����Ҷ�Ӷ࣬��������Ը߶��Եȸ߾Ӷࡣ���͡�Ҷ�Ρ�Ҷ����Ҷ���ܶȡ�Ҷ����ȡ�Ҷ�⼰Ҷɫ�����ȷֲ������������ϵ�2��3��������������ͱ���ΪС��ľ����ľ����С��ľΪ����Ҷ����ҪΪ��Բ�Ρ�����Բ�κ������Σ�������������Ҷ������Ϊ���ۺ�ƽ�������۾Ӷࣻ�~�����ܶ��ܡ�����оӶࣻҶ�����Ϊ����ͽ��⣬����ӶࣻҶɫ����ɫ������ɫ������ɫռ���������Ҷ�桢Ҷ�ʡ�Ҷ������Ƭ��ë����Ƭ��ɫ��������ɫ�������ʵء�������λ���ӷ���ë��Ҹѿ��ë����ѿ��ëȫ�����зֲ���1���������룬�������ΪҶ��¡��Ҷ���С�Ҷ��Ш�Ρ���Ƭ��ë�ޡ���Ƭ��ɫ��ɫ��������ɫ��ɫ�������ʵر���������λ�ߡ��ӷ�����ë��Ҹѿ�Ͷ�ѿ����ë��
9�������Ա�����״����Ƭ������ͷ���������ޱ仯���ȶ����ֳ���Ƭ5�����ͷ3�ѵ���״��Ҷ�������ͻ�����ͬ�������С��Ҷ���������ֳ�8�Ի�9�ԣ�����7��ӶࣻҶ����Ҷ����Ҷ���������ֱ�����������ȵı����Խϴ��Ŵ�������ָ��ͬ����������4����״��
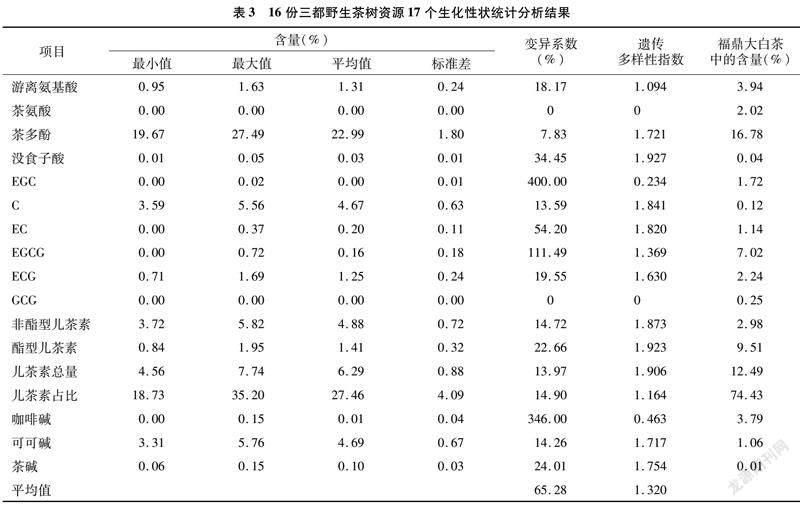
20�������Ա�����״����ϵ����0��35.68%֮�䣬ƽ��ֵΪ9.94%���Ŵ�������ָ����0��1.095֮�䣬ƽ��ֵΪ0.257��9�������Ա�����״����ϵ����0��32.60%֮�䣬ƽ��ֵΪ12.68%���Ŵ�������ָ����0��1.927֮�䣬ƽ��ֵΪ1.077��29��������״��ƽ������ϵ�����Ŵ�������ָ���ֱ�Ϊ10.79%��0.512����������Ұ����Դ������״�ı���ϵ�����Ŵ�������ָ�����ڽϵ�ˮƽ������11�������Ա�����״��2�������Ա�����״���зֲ�������ϵ�����Ŵ�������ָ����Ϊ0����˵������Ұ�������ڱ������ж����״����һ�£���Դ�ڲ������С������һ���ķ���ԡ�
2.2 ������״�Ŵ������Է���
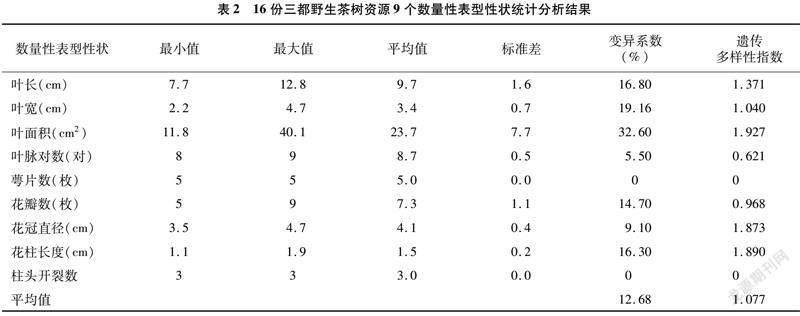
���ղ����г��������ɷֺ������Դ�����л����ı���16������Ұ��������17��������״���з������Ը�����ײ�Ϊ���ձȶ�������Դ���������������������״�ϵĵ��Ͳ��졣
�ɱ�3��֪��17��������״����ϵ�����Ŵ�������ָ������ֱ�Ϊ0��400.0%��0��1.927��ƽ��ֵ�ֱ�Ϊ65.28% ��1.320��������Դ�������ɷ�������ո�����ײ����ϴ�����ײ������백����Ͳ谱�Ậ���ֱ���3.94%��2.02%����������Դ�����백���Ậ���ϵͣ���߽�1.63%�����ڶ��ո�����ײ��һ�룬��16������Ұ�������о�δ���谱�ᡣ������ײ��в��Ӻ���Ϊ16.78%������������Ϊ12.49%��������ռ��Ϊ74.43%�����϶�����ռ���Ӻ���70.0%��80.0%�Ĺ��ɣ�15�ݣ���������Դ���ֳ����͵ĸ߲��ӡ� �Ͷ����ص����������Ӻ����������غ����Ͷ�����ռ��ƽ��ֵ�ֱ�Ϊ22.99%��6.29%��27.46%���Ա�����Ұ������������ײ�Ķ����ص����֪��������ײ�����Ҫ�����ص���Ϊ��ûʳ�Ӷ�����ûʳ��������EGCG��������Ϊ7.02%��ռ������������56.2%��������Դ��ռ�����Ķ����ص���Ϊ�����أ�C����������3.59%��5.56%֮�䣬����DZ�������ûʳ��������ECG����������0.71%��1.69%֮�䣬��ûʳ�Ӷ����أ�EGC����ûʳ�Ӷ�����ûʳ��������GCG����EGCG�ͱ������أ�EC���������ϵ͡��Ա��������ɿ�֪��������ײ�����Ҫ������ǿ��ȼ����Ϊ3.79%��������Դ�е���Ҫ�����Ϊ�ɿɼ������Χ��3.31%��5.76%֮�䣬�����������ȼ
2.3 �������������Է���
2.3.1 �������
����Դ����ڱ����22������ָ�꣨Ҷ����Ҷ����Ҷ�����Ҷ��������������������ֱ�����������ȡ������Ậ�������Ӻ�����ûʳ���Ậ����EGC������C������EC������EGCG������ECG�����������Ͷ����غ��������Ͷ����غ�����������������������ռ�ȡ����ȼ�����ɿɼ���Ͳ�����Ϊ���ݣ���16������Ұ���������о��������
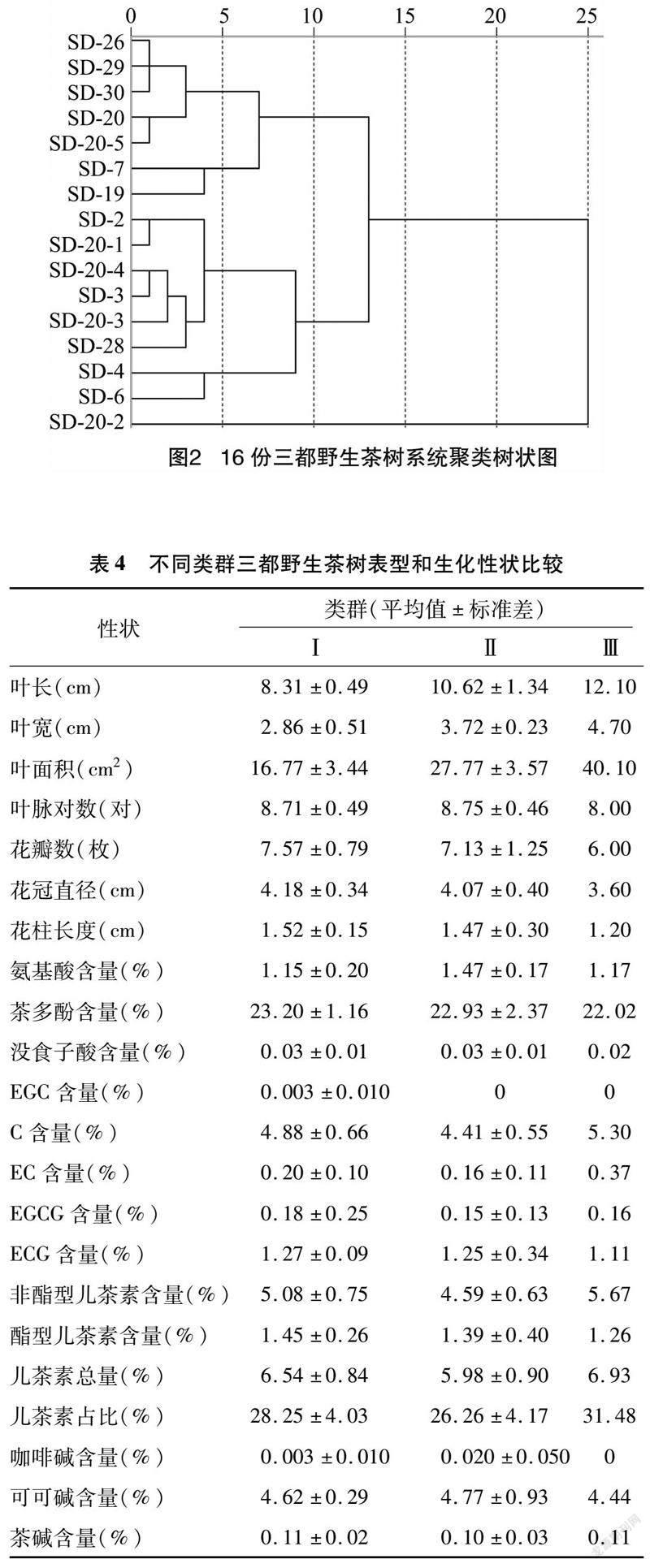
��ͼ2����4��֪��16������Ұ��������ŷʽ����Ϊ10ʱ���ɷ�Ϊ3�ࡣ�ڢ������SD-26��SD-29 ��7��Ұ����������Ҫ�ص�ΪҶ����Ҷ����Ҷ�������С�����Ӻ����Ͷ����غ����ϸߣ������Ậ���ϵͣ��ڢ������SD-2��SD-20-1��8��Ұ����������Ҫ�ص�Ϊ�����Ậ���ϸߣ�C�����������Ͷ����غ����Ͷ����������ϵͣ�������ռ�Ƚϵͣ��ɿɼ���ϸߣ�SD-20-2�������ɵڢ��࣬��Ҫ�ص�ΪҶ����Ҷ����Ҷ����ϴ��ӡ����Ͷ����غͿɿɼ���ϵͣ�C�������Ͷ����غ����Ͷ����������ϸߣ�������ռ�Ƚϸߡ�
2.3.2 ����Է���
ͬ������Դ����ڱ����22������ָ�����Pearson����Է������������5����ʾ������Ұ������������״��������״������Բ������ȼ��뻨����������������ء�������״��ҶƬ��״����Խ�ǿ��Ҷ����Ҷ����Ҷ���������ڼ���������أ�����״���л���ֱ���뻨�����ȼ���ڼ���������أ�ҶƬ��״�뻨��״�䲻������������ԡ�������״��������Ҫǿ�ڱ�����״����19��������״�������������������ԣ�ûʳ���Ậ����C��EC�� �����Ͷ����غ����������������Ͷ�����ռ�ȼ������������������ء�C������EC�������������ڼ���������أ�ͬʱ������Ͷ����غ����������������Ͷ�����ռ�ȼ������������������ع�ϵ��EGCG������ECG���������Ͷ����غ������ڼ���������ء�3�������ɷ��У��ɿɼ�����������ڼ���������ء�
3 ����
3.1 ����Ұ��������Դ�����Է���
����Ұ������������״�������ɷֵ��Ŵ�������ָ��ƽ��ֵ�ֱ�Ϊ0.512��1.320������������16�ݡ�������17�ݡ������18�ݼ����ݶ�صĵط��������ʣ�19-20�ݣ�Ҳ������Ӻ��21�ݡ�������22�ݡ��ײ���23�ݵȵص�Ұ��������˵������Ұ���������������Ϸ�գ����ͼ������Ŵ�������һ����Դ�ڲ��������Ͳ��ḻ��
3.2 ����Ұ��������Դ����
�ӷ�������ë���ӷ������Ͷ�ѿ��Ҹѿ���Ƿ��ë�Dz���ֲ������������Ҫ���ݣ�24�ݡ�����ֲ�C. sinensis���ļ�����������Ϊ�ӷ�3�ҡ���ë����ѿ��ë��25�ݣ�������Ұ�����������ӷ�3�ҡ���ë����ѿ��Ҹѿ����ë�ı����������ο������ֹ����Ž��衢ͻ�߲��Ԫ��ɽ����о���26�ݷ��֣�����Ұ����������̬�����Ž������ƶȽϸߣ���ͬ�����ӷ�3�ҡ���ë����ѿ��ë�⣬��Ҷ����Ҷ����Ҷ�����Ҷ�⡢Ҷ�Ρ������L�ȵ���״��ͬ������������������͡���Ƭ��ë����״�ϴ������Բ��죬�Ž���Ϊ��ľ����Ƭ�������ë������Ұ�����������ڹ�ľ��Ⱥ������Ƭ���⻬��ë�������ϣ�����Ұ���������Ž�������ֳ��Կɿɼ�Ϊ��Ҫ����CΪ��Ҫ�����ص��弰�谱�Ậ������������������ˣ���ϱ�����״���������ԣ��Ʋ�����Ұ��������Ԫ��ɽҰ������һ��������Ϊ�Ž�����֡�
3.3 ����Ұ��������Դ��������������
���ȼEGCG�Ͳ谱���Dz����е�������֣�15�ݡ������о����֣�Ұ���������������ڳ��������ֲ������������ԡ����ܵ��о����֣�ͺ���裨C. gymnogyna�����������������б����������֣��Կɿɼ�Ϊ��Ҫ������27�ݡ�Jin���о����֣�����Ұ����ѿ������Ҫ�����ص���ΪGCG����Ҫ���������ǿɿɼ��28�ݡ����������о����֣�����Ұ�������д����Կ���Ϊ��Ҫ�������������ʣ�29�ݡ�����Ұ��������Դ��C�Ϳɿɼ�Ϊ������֣��ɿɼ��֢����Ѫ�ܼ��������������ȷ����л�����Ԥ���������ã�30�ݣ���������Ϊ�������������Դ���Կ������á�
4 ����
����Ұ��������Դ���ͺ��������Ŵ�������ָ�������ڽϵ�ˮƽ���ڱ��ͺ���������ֻ��̶Ȳ��ߡ���Ҫ����������״����Ϊ�ӷ�3�ҡ���ë����ѿ��Ҹѿ����ë��������״������ֳ��߲��ӡ��Ͷ����ء��Ͱ�������谱����ص㣬C�Ϳɿɼ�������Ұ��������������������֡���ϱ�����״���������ԣ��Ʋ�����Ұ����������Ϊ�Ž�����֡����о���������Ұ��������Դ���ͼ���������������չ�о��������ɽ�Ϸǰ����л���������Ұ�������������ֵIJ����л�ͬʱ���ȫ���������ȷ����ֶΣ���������Ұ�������ڲ�����Դ�������еĵ�λ��
�ο����ף�
��1�����ľԣ��� ����Τ���죬��. ���������ѱ����Դ�봫����J��. ��������ԣ�2018��26��4����357-372.
��2�����DZ����Ի������Ǵ�ӣ���. Ұ��������������Դ��Ʒ��ָ������Է���������J��. ʳƷ��ҵ��2020��41��9����168-172.
��3��ţ���꣬�����������ڷɣ���. ����Ұ������������������״���������ۺ����ۣ�J��. �㽭ũҵѧ����2020��32��6����1039-1048.
��4���� �ƣ���Сë��Ф��������. ǧ��կҰ���Ų���ҶƬ���ʽṹ�ͻ�ѧ��ּ��������Ժ����ݶȵ���Ӧ��J��. ��̬ѧ��־��2021��40��7����1958-1968.
��5��Meegahakumbura M K��Wambulwa M C��Li M M��et al. Domestication origin and breeding history of the tea plant ��Camellia sinensis�� in China and India based on nuclear microsatellites and cpDNA sequence data��J��. Frontiers in Plant Science��2018��8��2270.
��6��Jiang C K��Zhao W F��Zeng Z��et al. A treasure reservoir of genetic resource of tea plant ��Camellia sinensis�� in Dayao Mountain��J��. Genetic Resources and Crop Evolution��2018��65��1����217-227.
��7��Du J K��He X L��Zhou Y M��et al. Gene coexpression network reveals insights into the origin and evolution of a theanine-associated regulatory module in non-Camellia and Camellia species��J��. Journal of Agricultural and Food Chemistry��2021��69��1����615-626.
��8�ݽ��ǿ���ų���������ǿ����. ����������Դ�о�“ʮ����”��չ��“ʮ����”��չ�����J��. �й���Ҷ��2021��43��9����42-49��76.
��9�ݳº���. ���ڻ������ز����ʾ���ݹŲ�����Դ��������о���D��. ���������ݴ�ѧ��2021��63-64.
��10�����࣬�Ե¸գ���ܲ�. �Ų���������Դ�Ŵ�������ISSR������J��. ���ӣ�2021��40��5����7-14.
��11�ݹ� �ӣ�Ƥ���꣬���������. ����GBS�����ȫ������SNP��ʾ���ݵط�����ֲ����Դ����Ե��ϵ��J��. �Ϸ�ũҵѧ����2021��52��3����660-670.
��12�ݾ���䣬���㷼��̷ �أ���. �������Բ�Ҷ���ʳɷֵ�Ӱ���J��. ʳƷ��ȫ�������ѧ����2015��6��4����1265-1270.
��13���� 漣��� ������ ������. ��ͬ����Ʒ�ְ�������ּ�����������J��. ����ũҵ��ѧ��2010��23����141-143.
��14����˳�䣬�� ѩ���ܹ�������. HPLC���ⶨ��Ҷ�ж����ؼ����ȼ��J��. ��ѧ����������2013��22��5����27-29.
��15������������ ��. ����������л��M��. ��������ѧ�����磬2015��1-3.
��16�ݶ�˧�Σ�����÷���� �ǣ���. ���ڱ�����״�������ɷֵ���������������Դ�Ŵ��������о���J��. ����ũҵѧ����2019��28��4����607-619.
��17�����λᣬ�� �������·��. ��������������ԴѿҶ���Ͷ����Է�����J��. ����ũҵ��ѧ��2020��48��1����134-138��142.
��18�ݵ��������� �⣬�� ������. ����30�ݲ���������Դũ����״�������ɷֶ����ԣ�J��. �㽭ũҵѧ����2021��33��7����1244-1255.
��19����С�٣��ܸ�ԣ���� ������. �������ֲ�����Դũ����״��Ʒ����״�����Է�����J��. ����ֲ�����֣�2015��13��2����415-423.
��20���� �����������ʯΰ������. ��ƽ�ط�������Դ�����ɷֶ����Է��������쵥���ѡ��J��. ���ӣ�2020��39��10����63-66��72.
��21�ݸ��������� ά������������. ���ݹ�ݼҰ��������̬�����Լ�������о���J��. ֲ���Ŵ���Դ�W����2015��16��3����497-502.
��22�ݲ� �꣬�Ǵ�ӣ��Ի�������. 25�ݹ�������Ұ������������Դ�ı��ͼ�������ֶ����Է�����J��. �й�ũѧͨ����2018��34��14����81-88.
��23���� �����������٣���. �ײ�Ұ��������Դ����������Ҫ�����ɷַ�����J��. ����ũҵѧ����2020��29��3����390-397.
��24�ݳ� �����ݸ�����ͯ����. ���ڲ���ֲ��������ݻ������ۣ�J��. ��Ҷ��ѧ��2000��20��2����89-94.
��25������». ����ɽ�������о���M��. ���������ϿƼ������磬2000��130.
��26�ݳ�����. ����Ԫ��ɽһ���������ֲ���ϵͳѧ�������ۺ������о���D��. ��ɳ������ũҵ��ѧ��2019��16-72.
��27���� �ܣ��� �꣬���ǻ�. ͺ�����������������ص㼰����Ʒ�ʳɷֵ��о���J��. ����ֲ�2018��38��5����568-576.
��28��Jin J Q��Chai Y F��Liu Y F��et al. Hongyacha��a naturally caffeine-free tea plant from Fujian��China��J��. Journal of Agricultural and Food Chemistry��2018��66��43����11311-11319.
��29�ݳ��������� �壬�����࣬��. ����Ұ��������Դ�������������ۼ�������Դɸѡ��J��. ��Ҷ��ѧ��2022��42��1����18-28.
��30����������ͯ ͮ���� ������. �ɿɲ輰�����ƻ�ѧ�ɷֵĽ�����Ч�о���չ��J��. ��Ҷ��ѧ��2021��41��5����593-607.